Recettori, Trasduzione del Segnale e Sistema del Complemento
Documento universitario sui recettori, la trasduzione del segnale e il sistema del complemento. Il Pdf esplora il riconoscimento dei patogeni, i meccanismi di difesa naturali e l'attivazione del complemento tramite via classica e alternativa, utile per Biologia.
Ver más12 páginas


Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Immunità e sistema del complemento
PROF
Reale Marcella
DATA
29/10/24
Si inizia con un ripasso della lezione precedente: La capacità del nostro sistema immunitario di difenderci.
Abbiamo detto che il primo step è quello di impedire l'ingresso del patogeno tramite le barriere che
ovviamente sono un meccanismo generico e passivo.
Nel caso in cui le barriere sono penetrate, quindi il patogeno arriva all'interno del nostro organismo, abbiamo
un meccanismo attivo; e quindi siamo già nella risposta che sappiamo essere inizialmente una risposta
naturale.
La risposta naturale è una prima risposta che entra in funzione quando siamo di fronte a un patogeno, ossia
ad un pericolo. Viene chiamata naturale perché è una risposta innata, nel senso che tutti noi ce l'abbiamo fin
dalla nascita. È naturale anche perché è uguale per tutti: lo stesso meccanismo è a disposizione di tutti noi ed
è lo stesso per qualunque patogeno incontriamo, quindi non è una risposta specifica, ma un meccanismo
unico, uguale per qualunque patogeno.
Affinché si abbia però una risposta, dobbiamo pero sapere nei confronti di chi dobbiamo rispondere. Quindi il
primo passo in questa risposta sarà il riconoscimento.
Le cellule coinvolte nella risposta immune naturale sono quelle dotate, nella maggior parte dei casi di attività
fagocitica. Per esempio:
- Granulociti in particolare neutrofili e dei macrofagi
- Cellule dendritiche
Riconoscimento patogeni e recettori
I granulociti e i macrofagi, per riconoscere patogeni, utilizzano
i recettori, che abbiamo definito come molecole PRR, cioè
recettori che riconoscono solo i profili molecolari associati al
patogeno oppure i profili molecolari associati al danno
cellulare.
I profili molecolari sono quelle molecole essenziali per la
sopravvivenza del patogeno, in quanto costituenti della
parete batterica e che sono costanti nel tempo; quindi, non
vanno incontro a modificazione e che quindi non sono invece
presenti nelle nostre cellule.
- Riconoscimento
C
KNIR
Riconoscimento
nel fagosoma
TLR
Riconoscimento
nel citosol
Recettore
per opsonine
PRR di
membrana
Opsonina
Riconoscimento
non opsonico
23/11/22Riconoscimento
opsonico
40
I recettori, situati sulle cellule fagocitarie, riconoscono qualcosa che non avevano mai visto prima che gli serve
a identificare la presenza del patogeno, ma non a specificare di quale patogeno stiamo parlando. Questo è un
riconoscimento di tipo naturale per differenziarlo poi da quello specifico.
Esistono delle molecole che servono a facilitare il riconoscimento e a instaurare dei legami aggiuntivi; tra la
cellula fagocitica e il patogeno che viene rivestito da queste molecole chiamate opsonine e di conseguenza si
avrà il riconoscimento del patogeno, e inizierà la trasduzione del messaggio.
Trasduzione del messaggio
La trasduzione del messaggio consiste in una riorganizzazione della cellula. Partendo dal citoscheletro, si ha
un'emissione di pseudopodi che poi si vanno a chiudere con il patogeno all'interno e quindi alla fine noi avremo
una vescicola delimitata da una membrana. Questo crea un ambiente isolato e protetto dal resto della cellula
per combattere il patogeno.
Questo è molto importante perché una volta che la cellula ha eliminato questo patogeno, ne può fagocitare un
altro.
Dal punto di vista dell'economia dell'individuo è estremamente importante che questa cellula non venga
danneggiata perché può diventare, un "individuatore" ed eliminatore del patogeno continuo.
Il fagosoma si deve fondere con il lisosoma, che contengono gli enzimi litici, i quali contribuiscono al
danneggiamento e quindi all'eliminazione del patogeno.
In alcuni casi non sono sufficienti e allora si attua un meccanismo di riserva.
È importante da ricordare che nella risposta immunitaria si parte sempre da un meccanismo più semplice e
economico dal punto di vista energetico per l'individuo e se questo non funziona abbiamo lo step successivo;
quindi, passiamo a qualcosa di più complesso e più impegnativo dal punto di vista energetico, ma non partiamo
subito da quello più impegnativo per una questione proprio di risparmio energetico per l'individuo.
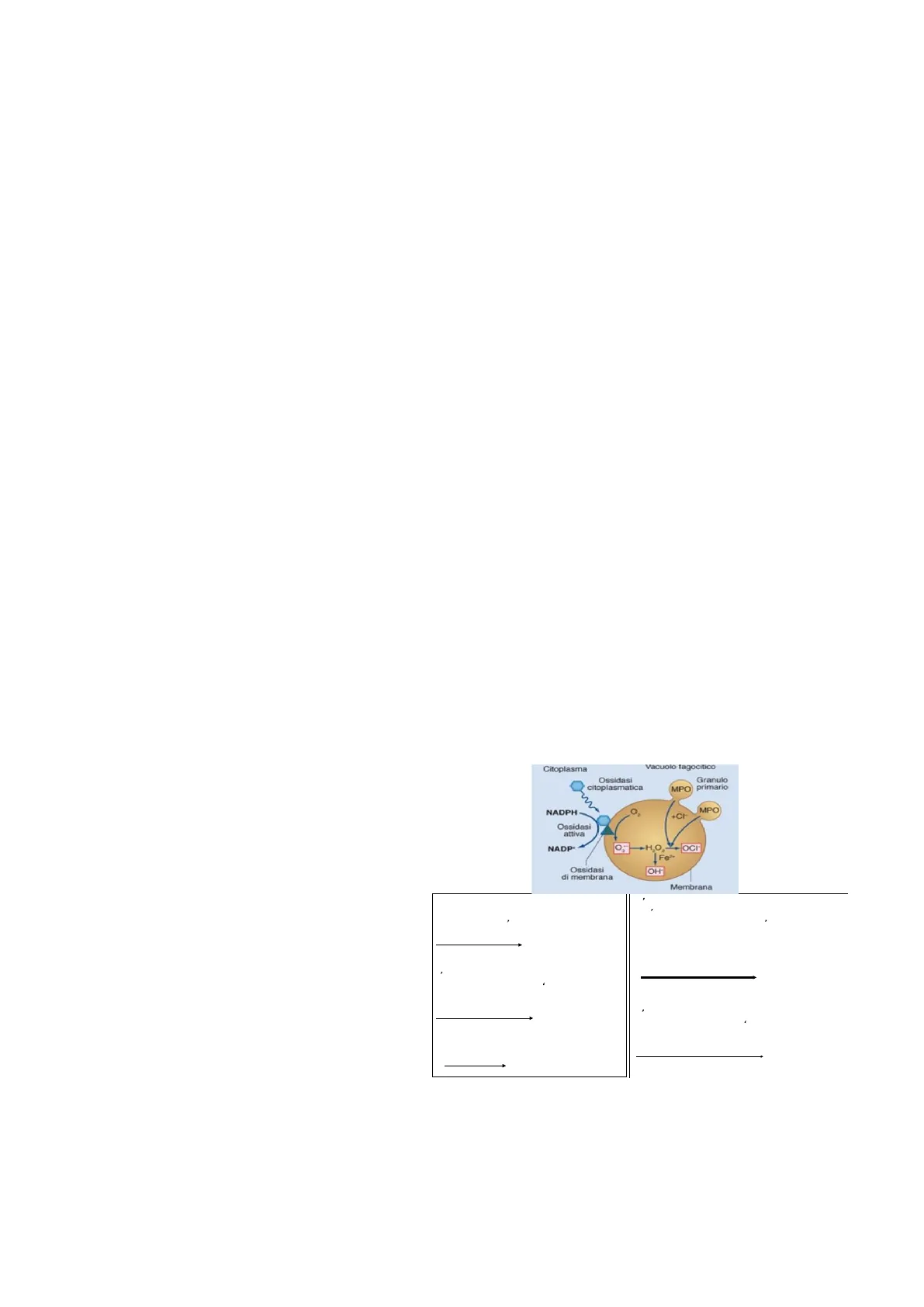
Se non sono sufficienti gli enzimi litici si ha l'attivazione di quella che è l'esplosione respiratoria ossidativa, che
porta alla produzione di specie reattive dell'ossigeno che come sono dannose per la cellula del dell'ospite, lo
sono anche per il patogeno, quindi vanno a danneggiare e contribuire all'eliminazione del patogeno.
Abbiamo detto che all'interno del fago lisosoma avviene la digestione del patogeno e distinguiamo i meccanismi
di distruzione di questo patogeno in meccanismi ossigeno indipendenti e ossigeno dipendenti.
I meccanismi ossigeno indipendenti sono appunto quelli a carico degli enzimi litici contenuti nel lisosoma,
mentre i meccanismi ossigeno dipendenti sono quelli che portano alla produzione delle specie reattive
dell'ossigeno, quest'ultimo si attiva solo dopo un certo lasso di tempo (30-40 secondi dall'inizio della fagocitosi)
e si attiva solo se è necessario, non parte subito, ma è in ritardo rispetto al meccanismo ossigeno indipendente
in quanto è dispendioso perché richiede un aumentato consumo di ossigeno, quindi una riduzione dell'ossigeno
molecolare disciolto nel sangue e nei liquidi
Citoplasma
Vacuolo fagocitico
interstiziali.
Importantissimo ricordare che è un meccanismo
che si attiva al bisogno, richiede l'attivazione
dell'ossidasi citoplasmatica che porta alla
produzione,
di
queste
specie
reattive
dell'ossigeno
con
un
meccanismo
energeticamente impegnativo per la cellula.
Possiamo dire che questo è il meccanismo di
Killing intracellulare.
Via
Ossigeno-dipendente
Ossidasi
citoplasmatica
Granulo
primario
MPO
Via
Ossigeno-dipendente
Reazioni
NADPH
O.
+CH
Reazioni
Ossidasi
attiva
Mieloperossidasi-
dipendenti
NADP.
0 +HOY.OCT
I Fe-
Ossidasi
di membrana
OH
Membrana
Il perossido di idrogeno reagendo con lo
ione cloro forma ione ipocloroso, la reazione
è catalizzata dall'enzima mielperossidasi
H2O2 + Cl-
OCI + H2O2
L'NADPH necessario per la riduzione
dell'ossigeno viene ottenuto per ossidazione del
glucoso-6-fosfato da parte dell'enzima glucoso-6-
fosfato deidrogenasi, quando diminuisce il rapporto
NADPH/NADP+
Glucoso 6-p +NADP+
G-6-P-dehydrogenase
6 fosfogluconato +
NADPH
L'anione superossido viene trasformato in
perossido di idrogeno dall ' enzima
superossido dismutasi
H2O2 + 02
L'anione superossido viene trasformato in
perossido di idrogeno dall ' enzima superossido
dismutasi
202 + 2H+
Superoxide dismutase
H2O2 + 102
H2O + O2 -
Mieloperossidasi
Indipendenti
Mielopessidasi
Abbiamo un'alternativa per quanto riguarda i
neutrofili nel caso in cui questo meccanismo non
2O2 - + 2H+
Superossido dismutasi
riesce ad eliminare il patogeno. Nel momento in
cui il patogeno non riesce ad essere fagocitato
Meccanismo protettivo
2H2O2
all'interno della cellula, ad esempio a causa delle
Catalasi
dimensioni superiori a quelle della cellula stessa, si ha un'alternativa.
I neutrofili riescono a formare i cosiddetti net, cioè delle trappole extra cellulari dove vengono intrappolati i
patogeni che sono costituite da sostanze battericide tipo elastasi neutrofila, gli istoni, mieloperossidasi,
lattoferrina e gelatinasi che vanno, a danneggiare il patogeno diminuendo le dimensioni che diventano più
accettabili e i detriti poi possono essere fagocitati.
2
MPCIl neutrofilo capisce che deve produrre questo net, grazie alle citochine pro-infiammatorie, le cellule endoteliali
attivate. Quindi abbiamo una serie di condizioni del microambiente e che consentono ai neutrofili di produrre
questi net. In definitiva, il neutrofilo
ha due possibilità
distinte
per
Meccanismo di rilascio NET
eliminare il patogeno: la fagocitosi,
bacteria
NET release
A
fungi
LPS
viruses
PMA
parasites
H.O.
Calcium ions
fMLP
IL-8
quindi eliminazione del patogeno per
C
Killing cellulare oppure la formazione
dei net, che portano alla distruzione e
ROS
bacteria
MPO
eliminazione del patogeno che viene
histones
prima degradato e poi i frammenti
B
vengono
invece
fagocitati dai
NE
chromatine
decondensation
NE
cell membrane rupture
neutrofili o da altri dai macrofagi ed
granule
MPO
PAD4
eliminati come frammenti, non più
come patogeno nel suo insieme.
(A) stimolazione dei recettori da parte di fattori scatenanti (es. batteri,
Si è visto, come le cellule della risposta
immune naturale, in particolare le
cellule
fagocitiche,
possono
contribuire
al mantenimento
del
nostro stato di salute, riconoscendo il
patogeno ed eliminandolo.
funghi, virus, parassiti, fattori chimici come PMA o LPS) porta all'adesione
dei neutrofili all'endotelio e alla decondensazione della cromatina dovuta
alla scissione dell'istone da parte di elastasi (NE) e mieloperossidasi (MPO)
e all'ipercitrullinazione dell'istone da PAD4 (B). La formazione di NET
richiede l'attivazione di NADPH ossidasi e produzione di ROS. Associati alle
fibre di NET ci sono lisozima, peptidi antimicrobici, componenti del
complemento e istoni.
Nella fase finale, i NET vengono rilasciati e intrappolano i batteri
53
Il meccanismo di eccellenza è la fagocitosi, quindi l'inglobamento all'interno della cellula e poi il Killing all'interno
del fago lisosoma che può essere ossigeno indipendente o ossigeno dipendente.
Nel caso in cui la fagocitosi non sia possibile, abbiamo un meccanismo alternativo a disposizione dei neutrofili,
ovvero la distruzione extracellulare del patogeno attraverso i net (successivamente si vedranno i macrofagi che
utilizzano le cellule specializzate).
Tuttavia, affinché questi meccanismi vadano avanti con successo abbiamo visto che è importante anche
l'opsonizzazione, cioè avere a disposizione molecole che rendano più "appetibile" il patogeno. Quando abbiamo
parlato della risposta immune naturale, abbiamo parlato anche dei mediatori solubili, quindi di quelle che sono
le "parole" e il "linguaggio" con cui le cellule comunicano. Dobbiamo quindi introdurre in questo discorso questi
mediatori, come lisozima e lattoferrina, che sono disponibili in ogni momento nel nostro organismo.
Alcuni mediatori sono più complessi e costituiscono un sistema e questi non sono immediatamente disponibili,
ma sono presenti nel nostro organismo in forma inattiva e quindi devono essere attivati. Sono inattivi perché
l'attivazione è un processo dispendioso e quindi che si attiva solo al bisogno.
Sistema del complemento
Tra questi, il sistema di mediatori più complessi è quello del complemento. Il complemento è cosi chiamato
perché va a complementare la risposta immune, sia quella naturale che quella specifica.
Questo sistema interviene sia nella risposta immune naturale sia nella risposta immune specifica,
complementando entrambe le risposte. È costituito da una serie di proteine sieriche termolabili.
Proteine sieriche; quindi, che le ritroviamo nel siero e termolabile, ossia che ad elevate temperature, in
particolare 56° si denaturano e quindi si perde l'attività di queste proteine.
Questa serie di proteine, attraverso una serie di passaggi ha come risultato finale l'induzione della morte
cellulare per lisi. Questa morte cellulare che può riguardare una cellula procariote o eucariote (quindi qualunque
cellula estranea di un altro individuo, una cellula batterica, una cellula trasformata, quindi tutto quello che può
essere considerato non desiderato dal nostro organismo, e che deve essere eliminato, ne provoca la morte per
lisi e questo è l'atto finale.
La peculiarità del sistema del complemento, che chiamiamo 'sistema' perché è costituito da più elementi, è che
produce, man mano che si attiva, mediatori intermedi che intervengono in altri momenti e in altri passaggi.
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
