Tema 2. La membrana plasmática: estructura, composición y funciones
Documento de universidad sobre Tema 2. La Membrana Plasmática. El Pdf, de Biología a nivel universitario, explora la morfología básica, la composición bioquímica y los mecanismos de adhesión y comunicación celular, incluyendo uniones intercelulares.
Ver más15 páginas


Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
La Membrana Plasmática
Es una "barrera" semipermeable que permite a las células compartimentalizar su citoplasma, delimitar su forma y comunicarse e interactuar con otras células. Además, permite el paso de sustancias del interior de la célula al exterior y viceversa. La membrana plasmática define el límite de las células, separando y manteniendo diferente concentración iónica entre el espacio extracelular y el intracelular, que es equivalente al citoplasma. El citoplasma está formado por el citosol, inclusiones y diferentes orgánulos, como la mitocondria o el aparato de Golgi. Algunos de estos orgánulos, como los ya citados o el aparato de Golgi, están delimitados por otras membranas intracelulares de composición similar a la membrana plasmática. Todas las membranas celulares comparten la característica de ser dinámicas y fluidas.
Morfología Básica de la Membrana
La membrana plasmática está formada por una estructura trilaminar de 8-10 nm de grosor, por lo que debemos recurrir a la microscopia electrónica. Si la observamos con muchos aumentos, observamos que esa estructura trilaminar está compuesta de dos bandas electrodensas del mismo tamaño y una banda electrolúcida. 2.5-3 nm 3.5-4 nm 2.5-3 nm En esta imagen se puede apreciar la estructura trilaminar de la membrana plasmática. Las dos bandas electrodensas corresponden a las cabezas polares de los fosfolípidos, y la banda electrolúcida equivaldría a las colas hidrofóbicas.
Revisión Histórica de la Membrana
- Los inicios de esta labor de investigación fueron los del trabajo realizado por Overton, hacia 1890. Él comprobó que mientras que las sustancias liposolubles penetraban en la célula con facilidad, las hidrosolubles apenas lo hacían. En 1895, presumió la existencia de una especie de cubierta celular que debía poseer naturaleza lipídica.
- Una década después, Langmuir estudió los lípidos de membrana como moléculas anfipáticas y propuso una organización en monocapa.
- En 1925, Gorter y Grendel extrajeron los lípidos de las membranas plasmáticas de un número conocido de eritrocitos y los extendieron en una monocapa. Este experimento condujo a la conclusión de que la membrana celular debía ser una bicapa lipídica.
- En 1930, gracias a la difracción de los rayos X se confirmó que la membrana era una bicapa lipídica. Sin embargo, eso no explico las funciones de las células, y se postulo que esas funciones las llevaban a cabo las proteínas
- En 1970, gracias a las micrografías TEM y a la criofractura se confirmó la existencia de proteínas transmembranas
-
Por último, en 1972, Singer y Nicholson propusieron el modelo "mosaico fluido", formado de fosfolípidos, colesterol y proteínas, mientras que el modelo anterior es de bicapa lipídica.
Modelo del Mosaico Fluido de Singer-Nicholson y Capaldi
Según el modelo de mosaico fluido, la estructura básica de la membrana es Proteína Proteina integral periférica Bicapa lipida una bicapa lipídica formada por lípidos anfipáticos en la que las porciones apolares de dichos lípidos se encuentran encaradas unas con otras en el pod Colesterol centro de la bicapa y sus grupos de cabeza polares encarados hacia el exterior a ambos lados de la misma. Las proteínas, que son de tipo globular, Fosfolípidos Glucolipido se encuentran incrustadas a intervalos irregulares en la bicapa Proteina Glucocalix periférica Glucoproteínas manteniendose unidas a ella mediante interacciones hidrofóbicas entre sus zonas apolares y las zonas apolares de los lípidos. La estructura es fluida, es decir, las moléculas individuales de lípidos y proteínas, debido a que se mantienen unidas por Proteína transmembranosa 1interacciones no covalentes, tienen libertad para moverse lateralmente en el plano de la membrana. La cara externa de la membrana plasmática, la que da al medio extracelular, presenta cadenas oligosacarídicas unidas covalentemente a lípidos o proteínas. Se trata de una estructura asimétrica por presentar diferencias entre su cara interna y su cara externa.
Composición Bioquímica de la Membrana
La membrana plasmática está formada por proteínas (45-50%) y por lípidos (50-55%). Las proteínas tienen función transportadora y receptora. Además, las glicoproteínas tienen función de reconocimiento. Dentro de los lípidos, hay tres tipos de moléculas lipídicas de membrana: fosfolípidos, Ácidos grasos colesterol y glucolípidos.
Fosfolípidos de la Membrana
- Fosfolípidos: dentro de los lípidos se corresponde con el 55%. Tienen una función de permeabilidad y de dar forma a la membrana. Podemos diferenciar ácidos neutros, como fosfatidiletanolamina (cefalina), fosfatidilcolina (lecitina) o esfingomielina; o fosfolípidos ácidos como fosfatidilinositol, fosfatidilglicerol, cardiolipina (mitocondrias), sulfolípidos o fosfatidilserina. Los fosfolípidos son moléculas anfipáticas que espontaneamente forman bicapas en las que los radicales polares quedan en contacto con el medio acuoso interno y externo.
-
Polo hidrofílico: es la cabeza polar y la parte electrodensa de la bicapa. Formada por glicerol, fosfato y un grupo polar como la colina (fosfatidilcolina o esfingomielina), etanolamida o serina.
-
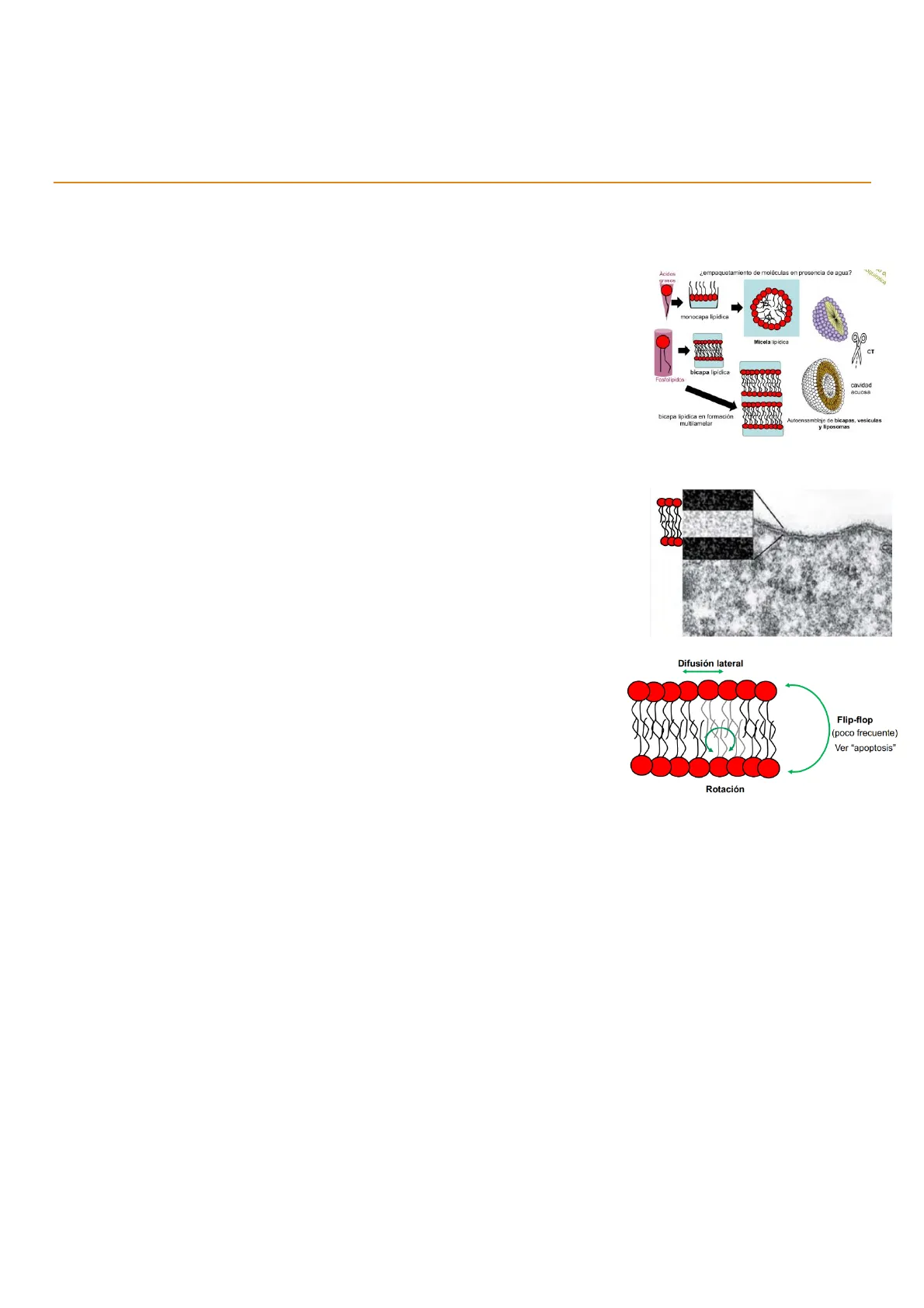
Polo hidrofóbico: es la cola de cadenas hidrocarbonadas (ácido graso de dos cadenas). Se corresponde con la parte electrolúcida. Eso hace que se distinga tridimensionalmente de los ácidos grasos u otros lípidos. Los ácidos grasos, al ocupar tridimensionalmente un espacio piramidal, forman una monocapa lipídica y una micela, mientras que los lípidos ocupan un espacio cilíndrico El grosor de la membrana plasmática influye en la fluidez de esta. Si la cola de los fosfolípidos está formada por ácidos grasos insaturados, tendrá menos grosor y por tanto tendrá más fluidez. Por el contrario, si los ácidos grasos fueran saturados, al aumentar el grosor disminuiría la fluidez de la membrana. ¿empaquetamiento de moléculas en presencia de agua? uquímica monocapa lipidica Micela lipidica CT bicapa lipidica Fosfolipidos cavidad acuosa bicapa lipidica en formación multilamelar Autoensamblaje de bicapas, vesiculas y liposomas Difusión lateral Flip-flop (poco frecuente) Ver "apoptosis" Rotación Por otro lado, los fosfolípidos son capaces de realizar ciertos movimientos que contribuyen a la fluidez de la membrana. El movimiento de difusión lateral es el más frecuente, y consiste en que los fosfolípidos pueden desplazarse lateralmente dentro de la bicapa. También pueden realizar una rotación sobre su eje longitudinal. El movimiento menos frecuente es el de flip-flop. Se trata del intercambio de fosfolípidos de una hemimembrana a la otra. Energéticamente es desfavorable debido a la dificultad que posee la cabeza polar para atravesar el medio hidrofóbico de la matriz de la membrana.
Colesterol y Glicolípidos
- Colesterol: se corresponde al 25% en el porcentaje de los lípidos. Su función principal es aportar rigidez y estabilidad a la membrana plasmatica. A mayor concentración de colesterol, menor fluidez de la membrana. Sin embargo, a menor temperatura menor fluidez de la membrana plasmática, siendo más fluida a mayor concentración de colesterol.
- Glicolípidos: cumplen con una función de reconocimiento celular. Están formados por un grupo glucídico (HdC) que se proyecta al espacio extracelular independientemente del número y la longitud de la cadena, y por un lípido que se encuentra insertado en la monocapa. Los glicolípidos se hallan involucrados en funciones de protección, de aislamiento y de reconocimiento celular, ya que conforman el glicocálix junto con las proteínas. 2Los glicolípidos están formados por una esfingosina, un ácido graso y un glúcido de cadena corta; y no tienen grupo fosfato. Dentro de los glicolípidos, en función del glúcido que forme la molécula, podemos distinguir:
Tipos de Glicolípidos
- Cerebrosidos: se encuentran en la membrana plasmática del sistema nervioso.
- Globósidos: es el antígeno P del eritrocito.
-
Gangliosidos: son los más complejos. Poseen ácido siálico, que tiene carga negativa, y se encuentra en las terminaciones nerviosas del SNC. Además, se encargan del sistema AB0 de la sangre. superficie extracelular Complejo de la union Espectrina Acting citoplasma Proteinas de unión Proteinas transmembrana 100 nm En cuanto a las proteínas, estas están inmersas dentro de la bicapa de los lípidos (proteínas integrales o intrínsecas). Algunas de las proteínas Alberts et al. 2014 ocupan todo el grosor de la membrana (proteínas transmembrana), quedando expuestas en ambas superficies, mientras que otras están inmersas en la hoja interna o externa de lípidos, sobre todo en la hemimembrana P. Las regiones de estas proteínas que sobresalen de la bicapa lipídica son hidrófilas. Otras proteínas de la membrana se encuentran ancladas a estructuras citoplasmáticas a través del citoesqueleto. Las proteínas periféricas de la membrana están unidas a la hoja interna o externa de la misma mediante enlaces no covalentes débiles con otras proteínas o lípidos. Las proteínas de la membrana son importantes en la adherencia entre las células y la matriz, la transmisión de señales intercelulares y la formación de canales transmembrana para permitir la entrada y salida de sustancias de la célula. Las características de las proteínas de membrana son las siguientes:
Características de las Proteínas de Membrana
- Realizan movimientos de rotación.
- Realizan la difusión lateral.
- No hacen cambios por flip-flop.
- Hay asimetría tanto entre hojas como en una misma hoja, en lo que se conocen como dominios de membrana, que contribuyen a la polarización.
El Glucocálix
En la superficie externa de las membranas plasmáticas de las células animales, la mayor parte de las proteínas y parte de los lípidos de la membrana se conjugan con cadenas cortas de polisacáridos. Estas glucoproteínas y glucolípidos se proyectan a partir de la superficie de la bicapa formando el glucocálix, cuyo grosor es variable según el tipo de célula (contribuye a la asimetría de la membrana). El glucocalix interviene en fenómenos de reconocimiento celular, en la formación de las adherencias intercelulares y en la adsorción de moléculas a la superficie celular. En ocasiones, también proporciona protección mecánica y química. Las microvellosidades están cubiertas por glicocálix, éste posee enzimas que participan en las últimas etapas de degradación de polisacáridos y proteínas. (+mecanismos de absorción en la membrana). glycocalyx cytosol nucleus plasma membrane 200 nm La cubierta celular o glicocalix de un linfocito teñida con rojo rutenio. Se marca la gruesa capa de carbohidratos que tapiza a la célula. 3
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
