La membrana plasmatica: composizione, funzioni e modello a mosaico fluido
Documento sulla membrana plasmatica, composizione e modello a mosaico fluido. Il Pdf esplora i ruoli di lipidi, proteine e carboidrati, la fluidità della membrana e l'influenza del colesterolo, utile per lo studio universitario di Biologia.
Mostra di più15 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
La Membrana Plasmatica: Struttura e Funzionamento
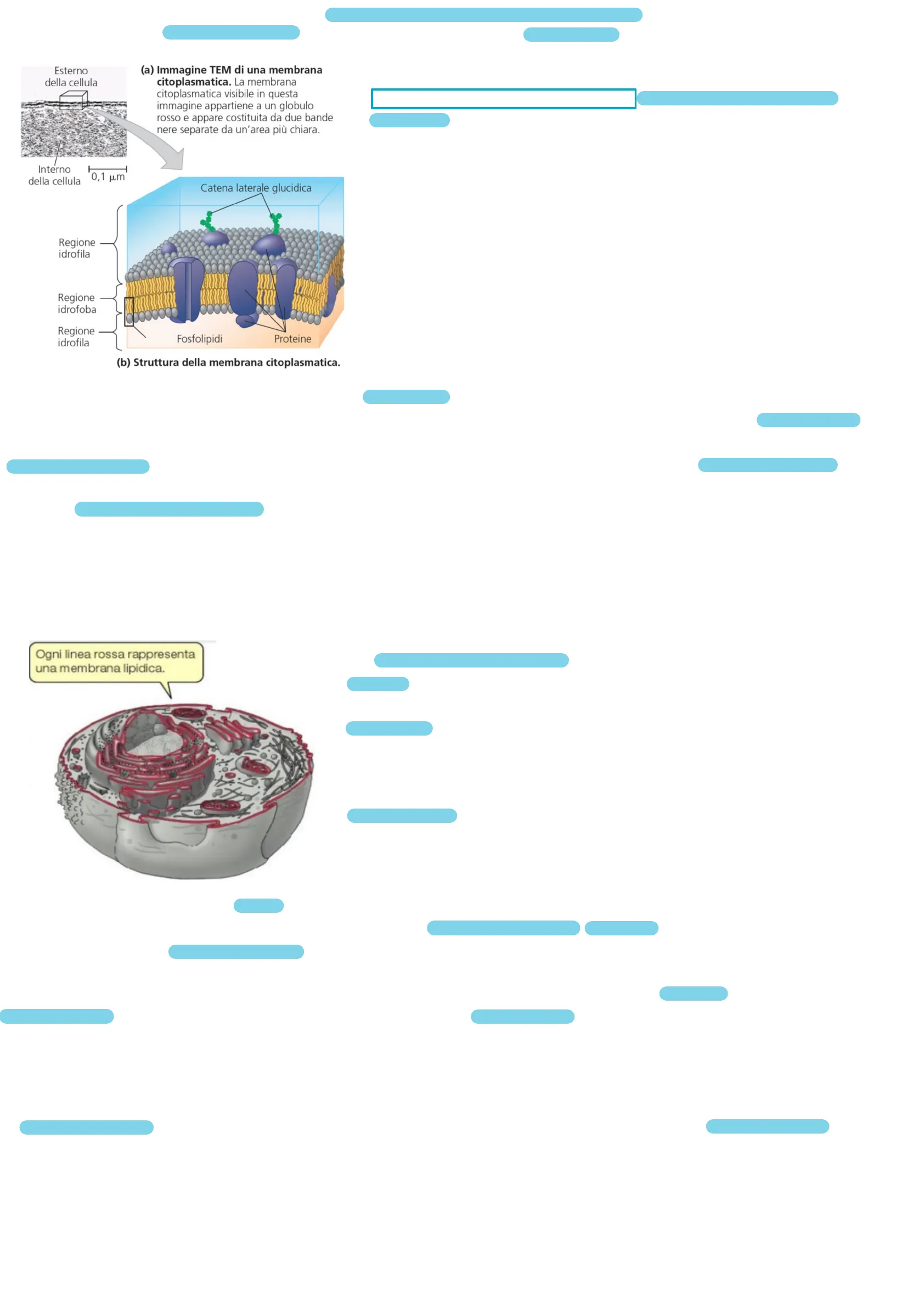
1La MEMBRANA PLASMATICA ha una struttura e un funzionamento unitario in tutti gli esseri viventi, formata principalmente da lipidi e proteine e in piccola percentuale da carboidrati. Esterno della cellula (a) Immagine TEM di una membrana citoplasmatica. La membrana citoplasmatica visibile in questa immagine appartiene a un globulo rosso e appare costituita da due bande nere separate da un'area più chiara. Interno della cellula 0,1 pm Catena laterale glucidica Regione idrofila Regione idrofoba Regione idrofila Fosfolipidi Proteine (b) Struttura della membrana citoplasmatica.
S.J Singer e G.Nicolson (1972): MODELLO A MOSAICO FLUIDO. Il modello ipotizza una membrana come un mosaico di proteine incluse in modo discontinuo in un doppio strato lipidico fluido
La membrana è formata principalmente da fosfolipidi (ad eccezione degli archeobatteri) che si organizzano a formare un doppio strato, siccome la membrana si posizione in un ambiente acquoso, le teste polari dei fosfolipidi formano legami a idrogeno con l'ambiente acquoso interno ed esterno alla cellula, mentre le code idrofobiche interagiscono tra loro. Quindi i fosfolipidi hanno principalmente un ruolo strutturale, determinano la struttura portante della barriera che circonda la cellula dall'ambiente esterno
C'è una componente proteica. Le proteine si dispongono con diversi rapporti con i fosfolipidi. Le proteine svoglono tutte le funzioni attive della membrana
I carboidrati sono presenti principalmente negli eucarioti e sono presenti principalmente legati ad altre molecole, a proteine sottoforma di glicoproteine, in alcuni casi sono legati a lipidi e la parte oligosaccaridica è rivolta verso l'esterno sempre
Ogni linea rossa rappresenta una membrana lipidica.
Componenti delle Membrane Biologiche
Le membrane biologiche sono composte da:
- Lipidi:
- Barriera idrofobica
- Proteine:
- Trasporto Specifico
- Riconoscimento e comunicazione
- Conversione di energia
- Carboidrati:
- Riconoscimento e comunicazione
Tutti i lipidi di membrana sono ANFIPATICI
Modello a Mosaico Fluido
In mosaico fluido, Il termine fluido indica che la membrana è fatta da fosfolipidi in quanto sono costituiti da acidi grassi che ha temperature normali hanno una consistenza fluida, mosaico perché le proteine si possono disporre in maniera sparsa, asimmetrica, non ordinata e si possono muovere tranquillamente, grazie appunto a questa fluidità.
All'interno della membrana a seconda delle nostre esigenze si possono inserire steroidi. Negli animali all'interno della membrana plasmatica c'è il colesterolo. Si posiziona principalmente nelle nostre membrane per regolarne lo stato fisico. È un tampone che permette alle nostre membrane di mantenersi in equilibrio di densità, né troppo fluida né troppo rigida, la giusta consistenza è fondamentale in modo che la membrana possa funzionare (se è troppo rigida le proteine rimangono bloccate e non riescono a muoversi, se troppo fluida le proteine rischiano di uscire dalla membrana)
le cellule vegetali non hanno il colesterolo, ma delle molecole prodotte da loro stessi, FITOSTEROLI, che hanno una struttura simile al colesterolo e svolgono le stesse funzioni(a) FOSFOLIPIDI Fosfoglicerolipidi (fosfogliceridi) Fosfatidilcolina (mostrato) Fosfatidiletanolamina Fosfatidilserina Fosfatidiltreonina Fosfatidilinositolo Fosfatidilglicerolo Difosfatidilglicerolo (cardiolipina) Fosfosfiingolipidi Colina Sfingomielina (uno sfingolipide) Colina Fosfato Fosfato 1 2 2 Glicerolo Sfingosina Acido grasso Acido grasso Acido grasso (b) GLICOLIPIDI Glicoglicerolipidi Glicosfingolipidi Cerebrosidi Monogalattosildiacilglicerolo (MGDG, mostrato) Galattosio Galattosio Digalattosildiacilglicerolo (DGDG) (galattocerebroside, mostrato) 2 1 2 العيا Glicerolo Sfingosina Acido grasso Acido grasso Acido grasso (c) STEROLI Steroli Colesterolo (mostrato) Campesterolo Sitosterolo Fitosteroli
La membrana plasmatica degli eubatteri ha degli steroli con la stessa funzione, hanno molecole molto simili al colesterolo chiamate OPANOIDI
Il rapporto degli elementi che costituiscono le membrane varia in base alle esigenze.
Composizione in Fosfolipidi di Diverse Membrane
Membrana plasmatica (fegato di ratto) Membrana plasmatica (patata) Membrana plasmatica (E. coli) Guaina mielinica (umana) Membrana tilacoide del cloroplasto (spinaci) Membrana Percentuale totale di fosfolipidi 60 40 20 0 Fosfatidiletanolamina Fosfatidilinositolo Fosfatidilcolina Fosfatidilglicerolo Fosfatidilserina Difosfatidilglicerolo (cardiolipina) Sfingomielina FIGURA 7.7 Composizione in fosfolipidi di diversi tipi di membrane. L'abbondanza relativa dei diversi tipi di fosfolipidi nelle membrane biologiche varia moltissimo in relazione alla provenienza della membrana.
Funzioni delle Proteine di Membrana
FUNZIONI DELLE PROTEINE DI MEMBRANA: (a) Trasporto. A sinistra: una proteina che attra- versa la membrana a tutto spessore può for- mare un canale idrofilo che consente il pas- saggio selettivo di un soluto specifico. A destra: altre proteine di trasporto trasferi- scono una sostanza da un lato all'altro grazie a un cambiamento conformazionale (si veda la Figura 7.14b). Alcune di queste idrolizzano l'ATP al fine di ottenere l'energia necessaria per pompare attivamente le sostanze attraverso la membrana. (b) Attività enzimatica. Una proteina di mem- brana può svolgere una funzione enzimatica, con il sito attivo esposto alle sostanze pre- senti nell'ambiente circostante. In alcuni casi, gli enzimi di membrana sono organizzati all'interno di un complesso in grado di cata- lizzare tappe successive di una specifica via metabolica. (c) Trasduzione del segnale. Una proteina di membrana (recettore) può presentare un sito di legame specifico per un messagge- ro chimico, per esempio un ormone. Il legame del messaggero esterno (molecola segnale) potrebbe determinare una modificazione conformazionale della proteina, consentendo il trasferimento del messaggio all'interno della cellula, in genere mediante il legame di una proteina citoplasmatica (si veda la Figura 10.6). (d) Riconoscimento fra cellule. Alcune glico- proteine agiscono da marcatori che vengono riconosciuti specificamente da proteine di membrana di altre cellule. Questo tipo di legame cellulare solitamente è di breve durata rispetto a quello illustrato in (e). Glico- proteina AT Enzimi (e) Adesione intercellulare. Le proteine di membrana di cellule adiacenti possono unirsi insieme per formare diversi tipi di giunzione, per esempio le giunzioni serrate e le giunzioni occludenti (si veda la Figura 6.30). Questo tipo di legame è di maggiore durata rispetto a quello mostrato in (d). Molecola segnale Recettore Trasduzione del segnale (f) Adesione al citoscheletro e alla matrice extracellulare. I microfilamenti o altri ele- menti del citoscheletro possono legarsi (in modo non covalente) alle proteine di membrana, fissando la posizione di queste e contribuendo al mantenimento della forma dell'intera cellula. Le proteine che si legano alle molecole della matrice extracellulare pos- sono coordinare cambiamenti extra- e intra- cellulari (si veda la Figura 6.28). FIGURA 7.6 Le tre principali classi di lipidi di membrana. (a) I fosfolipidi sono costituiti da una piccola testa polare (come la colina) attaccata tramite un gruppo fosfato a uno scheletro di glicerolo o sfingosina contenete acidi grassi, a formare o fosfoglicerolipidi (fosfogliceridi) o fosfosfingolipidi. Per il glicerolo e la sfingosina sono segnati gli atomi di carbonio 1, 2 e 3. (b) I glicolipidi hanno uno o più monosaccaridi ancorati a uno scheletro contenente acidi grassi e anch'essi possono essere a base o di glicerolo o di sfingosina. (c) Gli steroli sono molecole a più anelli che sono correlati al colesterolo e agli ormoni steroidei. Gangliosidi Stigmasterolo Ergosterolo Opanoidi mitocondriale interna (fegato di ratto)
Ricerca Scientifica e Membrana Plasmatica
Ricerca scientifica e membrana plasmatica:
1880 Apolare Polare (a) Natura lipidica della membrana Lipidi Overton 1900 (b) Monostrato lipidico Langmuir Irving Langmuir: Modello del Mono-strato lipidico. (c) Doppio strato lipidico -1920 Gorter. e Grendel (d) Doppio strato lipidico e strati proteic Davson e Danielli -1940 (e) Membrana unitaria Robertson 1960 (f) Modello a mosaico fluido Singer e Nicolson Unwin e Henderson Singer e Nicolson (1972): Modello a mosaico fluido. Nigel Unwin e Richard Henderson (1975): Alcune proteine intrinseche hanno più segmenti transmembrana Le membrane, modificando la loro composizione, possono formare microdomini chiamati zattere lipidiche (lipid raft)
Studi di Overton sulla Natura Lipidica
Overton (1890): natura lipidica della membrana Egli osservò che le sostanze solubili nei lipidi penetravano più facilmente di quelle solubili in acqua. Ipotizzò che i lipidi fossero presenti sulla superficie cellulare, come una sorta di «rivestimento». Overton fece una serie di esperimenti misurando la velocità di passaggio all'interno di una cellula di una serie di sostanze di varia natura, polare, idrofobica e anfipatica. Vide che le sostanze polari non erano in grado di passare, mentre erano in grado di passare quelle idrofobiche. Ipotizzò quindi che le membrane avessero una natura lipidica. Restava ora di capire come i lipidi si potessero organizzare per formare la membrana. Il primo a ipotizzarlo fu Irving Langmuir.
Modello del Monostrato Lipidico di Langmuir
Irving Laugmuir: modello del Mono-strato lipidico Dieci anni dopo gli studi di Irving Langmuir portarono alla prima ipotesi sulla struttura della membrana plasmatica. · Studiò il comportamento dei fosfolipidi purificati e disciolti in un solvente organico (benzene). · Ponendo la miscela di fosfolipidi in benzene in acqua, quando il benzene evaporava si formava un monostrato di fosfolipidi in cui la testa polare rimaneva immersa nell'acqua, mentre la porzione apolare si esponeva all'aria.
Modello del Doppio Strato Lipidico di Gorter e Grendel
Gorter e Grendel (1925): modello del doppio strato lipidico · Due fisiologi tedeschi, Gorter e Grendel, cercarono di scoprire quanti strati lipidici sono presenti nella membrana plasmatica. · Estrassero i fosfolipidi dagli eritrociti e utilizzando il metodo di Langmuir, stratificarono questi su una superficie acquosa. · Calcolarono l'area coperta dal film di fosfolipidi: risultò essere circa il doppio dell'area occupata dalla superficie del globulo rosso. · Dedussero che ogni globulo rosso doveva possedere due strati di fosfolipidi. ARIA lipidi gruppo fosforico ACQUA ACQUA GRUPPI FOSFORICI STRATO LIPIDICO GRUPPI FOSFORICI ACQUA Apolare- Polare - Natura lipidica della membrana Lipidi (h) Zattere lipidiche
Overton : Natura lipidica della membrana. Ipotizzò che i lipidi fossero presenti sull a superficie cellulare, come una sorta di «rivestimento».
Gorter e Grendel (1925): Modello del doppio strato lipidico.
Davson e Danielli (1935): Modello a sandwich.
Roberston (1950): grazie alla microscopia elettronica osservò che le cellule e le strutture interne erano circondate dalla stessa struttura che chiamò Membrana Unitaria.
1980 (g) Struttura di una proteina di membrana a elica 2000
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
