Glucólisis y Gluconeogénesis: Vías Metabólicas Centrales en Biología
Documento sobre Glucólisis y Gluconeogénesis. El Pdf detalla las reacciones químicas, fases y balance energético de estos procesos fundamentales en Biología, adecuado para el estudio universitario.
Ver más15 páginas


Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Bioquímica II. Grado en Biología
Tema 8.1. Glucolisis/gluconeogénesis
- Generalidades: Función biológica, localización subcelular, balance.
- Descripción de la ruta.
- Gluconeogénesis a partir de piruvato, lactato, aminoácidos, glicerol.
- Regulación.
- Incorporación de otros azúcares a la ruta.
- Bibliografía.
Generalidades de la Glucólisis
La glucosa ocupa una posición central en el metabolismo de plantas, animales y muchos microorganismos. Es utilizada como combustible, de manera que su oxidación puede abastecer de energía a las células. La glucolisis es la ruta por la que se degrada la glucosa hasta piruvato. En esta ruta confluye la degradación de otros carbohidratos como la fructosa, galactosa, manosa, sacarosa, lactosa, almidón y glucógeno, convirtiéndose por tanto en la ruta central de degradación de carbohidratos. Por otro lado, la glucosa es utilizada como fuente de carbono a partir de la cual pueden sintetizarse el resto de moléculas biológicas tales como aminoácidos, nucleótidos, coenzimas, ácidos grasos y otros intermediarios.
Es una ruta ubicua que ocurre en el citosol de todas las células, desde los organismos más sencillos a los más complicados.
La formulación de la ruta completa fue propuesta, en la década de los años 30 del siglo pasado, por Otto Warburg y Hans von Euler-Chelpin, en levaduras, y por Gustav Embden y Otto Meyerhof, en el músculo.
Otto Warburg, 1883-1970 Hans von Euler-Chelpin, 1873-1964 Gustav Embden, 1874-1933 Otto Meyerhof, 1884-1951
El balance global de la ruta es: Glc + 2 ADP + 2 Pi + 2 NAD+ -> 2 Pir + 2 ATP + 2 NADH + 2 H+ + 2 H2O, AG1º = - 85,0 KJ/mol (1)
Es una ruta única ya que puede funcionar en presencia como en ausencia de oxígeno. En condiciones aeróbicas la glucosa se degrada hasta 2 moléculas de piruvato generando 2 moléculas de ATP y 2 moléculas de NADH + H+. En estas condiciones, la glucolisis es la primera parte de la degradación completa la glucosa. El piruvato generado se transformará en acetil-CoA que pasa posteriormente por el ciclo de los ácidos tricarboxílicos y por la cadena mitocondrial de transporte de electrones para ser finalmente transformado en CO2, H2O y ATP. Pueden generarse hasta 32 moléculas de ATP por molécula de glucosa. En condiciones anaeróbicas se produce la transformación de glucosa hasta lactato o etanol con la producción de 2 moléculas de ATP por molécula de glucosa.
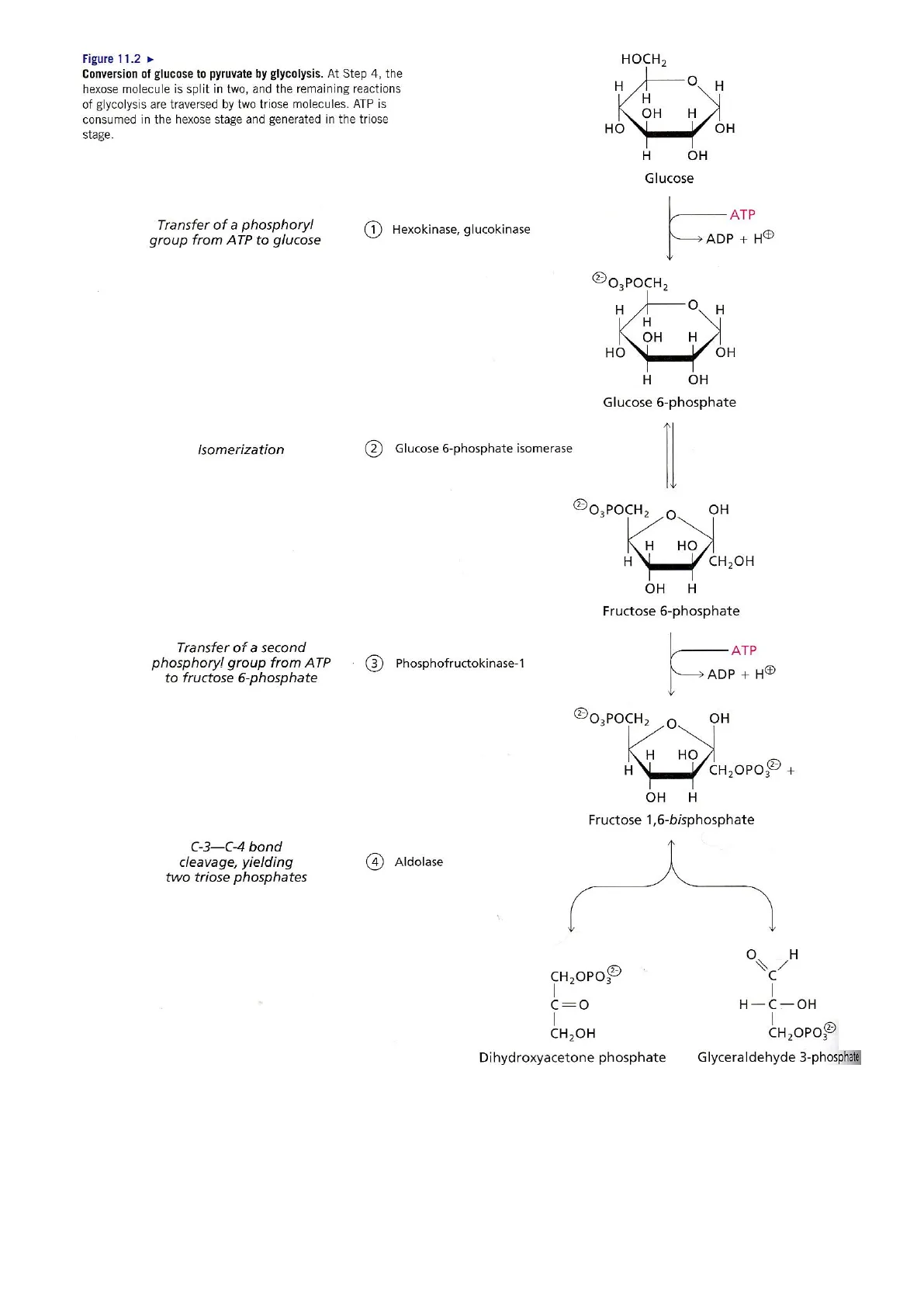
Glc + 2 ADP + 2 Pi -> 2 Lactato + 2 ATP + 2 H2O (2) 1Figure 11.2 Conversion of glucose to pyruvate by glycolysis. At Step 4, the hexose molecule is split in two, and the remaining reactions of glycolysis are traversed by two triose molecules. ATP is consumed in the hexose stage and generated in the triose stage. Transfer of a phosphoryl group from ATP to glucose 1 Hexokinase, glucokinase 2- O3 POCH2 O H H H OH H HO OH H OH Glucose 6-phosphate Isomerization 2 Glucose 6-phosphate isomerase 203 POCH 2 OH O H HO H CH2OH OH H Fructose 6-phosphate Transfer of a second phosphoryl group from ATP to fructose 6-phosphate 3 Phosphofructokinase-1 ATP > ADP + HO 2 O3 POCH2 O H HO H CH2OPO,3 + OH H Fructose 1,6-bisphosphate C-3-C-4 bond cleavage, yielding two triose phosphates 4 Aldolase HOCH 2 O H H H OH H HO OH H OH Glucose - ATP > ADP + HD CH2OH CH2OPO32 Dihydroxyacetone phosphate O H 11 CH2OPO3 1 v- C=0 H-C-OH Glyceraldehyde 3-phosphate Procedencia de la Ilustración: Moran y col. (2012). Principles of Biochemistry. Pearson. 2 OHBioquímica II. Grado en Biología Tema 8.1. Glucolisis/gluconeogénesis O H CH2OH c=0 - H-C-OH Triose phosphate isomerase 5 Rapid interconversion of triose phosphates CH2OPO3 2- Dihydroxyacetone phosphate Glyceraldehyde 3-phosphate CH2OPO 2 NAD+ Pi >NADH + HO Glyceraldehyde 3-phosphate dehydrogenase 6 Oxidation and phosphorylation, yielding a high-energy mixed-acid anhydride O OPO3 U H-C-OH CH2OPO3 2- 1,3-Bisphosphoglycerate ADP Phosphoglycerate kinase 7 Transfer of a high-energy phosphoryl group to ADP, yielding ATP H-C-OH - CH2OPOS 3-Phosphoglycerate Phosphoglycerate mutase 8 Intramolecular phosphoryl-group transfer COOO H-C-OPO3 CH 2OH 2-Phosphoglycerate H20 1 > H2O Enolase 9 Dehydration to an energy-rich enol ester Phosphoenolpyruvate COOP -OPOR II CH2 k ADP + HO > ATP Pyruvate kinase 10 Transfer of a high-energy phosphoryl group to ADP, yielding ATP COOP C=0 I CH3 Pyruvate 3 12 > ATP
Descripción de la Ruta Glucolítica
Es una ruta constituida por diez reacciones químicas catalizadas por enzimas citosólicos en la que se pueden distinguir dos fases: la fase de las hexosas y la fase de las triosas.
Fase de las Hexosas
a) Fase de las hexosas: en la que la glucosa se prepara para su catabolismo. Consiste en la fosforilación de ésta y su transformación hasta fructosa 1,6-bisfosfato. En esta fase se consumen 2 ATP por cada molécula de glucosa.
Fase de las Triosas
b) Fase de las triosas: Las triosas fosfato obtenidas por rotura aldólica de la F16BP se convierten en piruvato produciéndose reacciones de oxidación-reducción y formación de ATP. En esta fase se forman 4 ATP por cada molécula de glucosa.
Reacciones de la Fase de las Hexosas
Reacción 1: La glucosa (Glc) se transforma en glucosa 6-fosfato (G6P) mediante una reacción de transferencia de grupo fosforilo (fosforilación) catalizada por la hexoquinasa (músculo) o glucoquinasa (hígado). El grupo y-fosforilo del ATP es transferido hasta el átomo de oxígeno del carbono 6 de la glucosa.
Reacción 2: La glucosa 6-fosfato se transforma en fructosa 6-fosfato (F6P) mediante una reacción de isomerización catalizada por la glucosa 6-fosfato isomerasa. La reacción consiste en la transformación de una aldohexosa en una cetohexosa.
Reacción 3. La fructosa 6-fosfato se transforma en fructosa 1,6-bisfosfato (F16BP) mediante una reacción de fosforilación catalizada por la fosfofructoquinasa-1 (PFK-1). Es una reacción de transferencia del grupo y-fosforilo desde el ATP hasta el átomo de oxígeno del carbono 1 de la fructosa 6-fosfato. El sustrato de la reacción es el isómero ß-D-fructosa 6-fosfato.
Reacción 4. La fructosa 1,6-bisfosfato se transforma en gliceraldehido 3-fosfato (GADP) y dihidroxiacetona fosfato (DHAP) mediante una reacción de ruptura aldólica del enlace C-C existente entre el carbono 3 y carbono 4 de la F16BP catalizada por la enzima aldolasa.
Reacciones de la Fase de las Triosas
Reacción 5. La dihidroxiacetona fosfato se transforma en gliceraldehido 3-fosfato mediante una reacción de isomerización catalizada por la triosa fosfato isomerasa. La reacción consiste en la transformación de una cetohexosa en una aldohexosa. A partir de aquí todas las reacciones deben considerarse por duplicado para calcular la estequiometria global del proceso ya que por cada molécula de glucosa que entra en la ruta se producen 2 moléculas de GADP.
Reacción 6. El gliceraldehido 3-fosfato se transforma en 1,3-bisfosfoglicerato (1,3BPG) mediante una reacción de oxidación y fosforilación catalizada por la gliceraldehido 3-fosfato deshidrogenasa (GADPDH). En esta reacción se produce la reducción de 1 molécula de NAD+ que se transforma en 1 molécula de NADH + H+. 1,3BPG tiene un alto potencial de transferencia de grupos fosfato, mayor que el ATP. Esta es la reacción clave de la glucolisis ya que todas las reacciones anteriores son preparatorias para que ocurra ésta y las fases posteriores no son más que una consecuencia lógica (recogida de beneficios) de la creación de un compuesto con tan alta capacidad de transferencia de grupos.
Reacción 7. El 1,3-bisfosfoglicerato se transforma en 3-fosfoglicerato (3PG) mediante una reacción de fosforilación a nivel de substrato catalizada por la fosfoglicerato quinasa. Se transfiere el grupo fosfato del carbono 1 del 1,3BPG hasta el ADP generando ATP y 3PG.
Reacción 8. El 3-fosfoglicerato se transforma en 2-fosfoglicerato (2PG) mediante una reacción de isomerización catalizada por la fosfoglicerato mutasa. Las mutasas transfieren grupos fosforilo desde una parte a otra de una misma molécula.
4Bioquímica II. Grado en Biología Tema 8.1. Glucolisis/gluconeogénesis
Reacción 9. El 2-fosfoglicerato se transforma en fosfoenolpiruvato (PEP) mediante una reacción de deshidratación catalizada por la enolasa.
Reacción 10. El fosfoenolpiruvato se transforma en piruvato (Pir) mediante una reacción de fosforilación a nivel de substrato, en la que se genera una molécula de ATP, catalizada por la piruvato quinasa.
En condiciones celulares, 3 de estas reacciones son prácticamente irreversibles (1,3,10) y 7 son prácticamente reversibles tal y como se deduce de los cambios en la energía libre de cada una de ellas reflejados en la siguiente figura.
Figure 11.12 Cumulative standard and actual free-energy changes for the reactions of glycolysis. The vertical axis indicates free-energy changes in kJ mol-1. The reactions of glycolysis are plot- ted in sequence horizontally. The upper plot (red) tracks the standard free-energy changes, and the bottom plot (blue) shows actual free-energy changes in erythrocytes. The interconversion reaction catalyzed by triose phosphate isomerase (Reaction 5) is not shown. [Adapted from Hamori, E. (1975). Illustration of free energy changes in chemi- cal reactions. J. Chem. Ed. 52:370-373.] 0 - O -50 - 0 - -50 - Actual free-energy changes AG = - 72.3 KJ mol-1 -100 - 1 2 4 6 7 8 9 10 Reactions of glycolysis 3. Gluconeogénesis a partir de piruvato, lactato, aminoácidos, glicerol y propionato
Gluconeogénesis a partir de Piruvato, Lactato, Aminoácidos, Glicerol y Propionato
En muchos organismos (animales, plantas, hongos y microorganismos) existe una ruta de síntesis de glucosa a partir de intermediarios no glucídicos denominada gluconeogénesis. En el hombre, después de un ayuno de entre 16 y 24 horas, las reservas de glucógeno hepático se agotan y, ciertos tejidos como el hígado y el riñón -en menor medida-, pueden sintetizar glucosa a partir de precursores no glucídicos tales como el lactato y la alanina. Esta glucosa es imprescindible para que puedan sobrevivir células que dependen casi exclusivamente de ella, como los eritrocitos o las neuronas.
Gluconeogénesis a partir de Piruvato
La gluconeogénesis a partir de piruvato aprovecha las siete reacciones reversibles de la glucolisis y busca tres rodeos para realizar en sentido inverso los pasos irreversibles. Rodeo 1. Transformación de piruvato en fosfoenolpiruvato. El piruvato se transforma inicialmente en oxalacetato mediante una reacción de carboxilación catalizada por la piruvato carboxilasa, una enzima mitocondrial. En esta reacción se gasta una molécula de ATP y la biotina participa como grupo prostético transportador de CO2. La piruvato carboxilasa es una enzima mitocondrial, de forma que el piruvato debe entrar al interior de la mitocondria atravesando la membrana mitocondrial externa (porinas) y la membrana mitocondrial interna (piruvato translocasa). No existe un transportador de oxalacetato que pueda sacarlo fuera de la mitocondria directamente, de forma que es necesario dar un rodeo que consiste en la transformación de oxalacetato en malato, 5 Standard free-energy changes Cumulative free-energy changes (kJ mol-1) -O
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
