L'apoptosi cellulare: geni chiave, caspasi e vie di attivazione
Documento universitario sull'apoptosi cellulare. Il Pdf esplora i meccanismi molecolari che la regolano, inclusi i geni ced, le caspasi e le vie intrinseca ed estrinseca, per la materia Biologia.
Mostra di più12 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Biologia II
13/05/2024
Guglielmo Pizzigallo
Francesca Paris
L'apoptosi cellulare
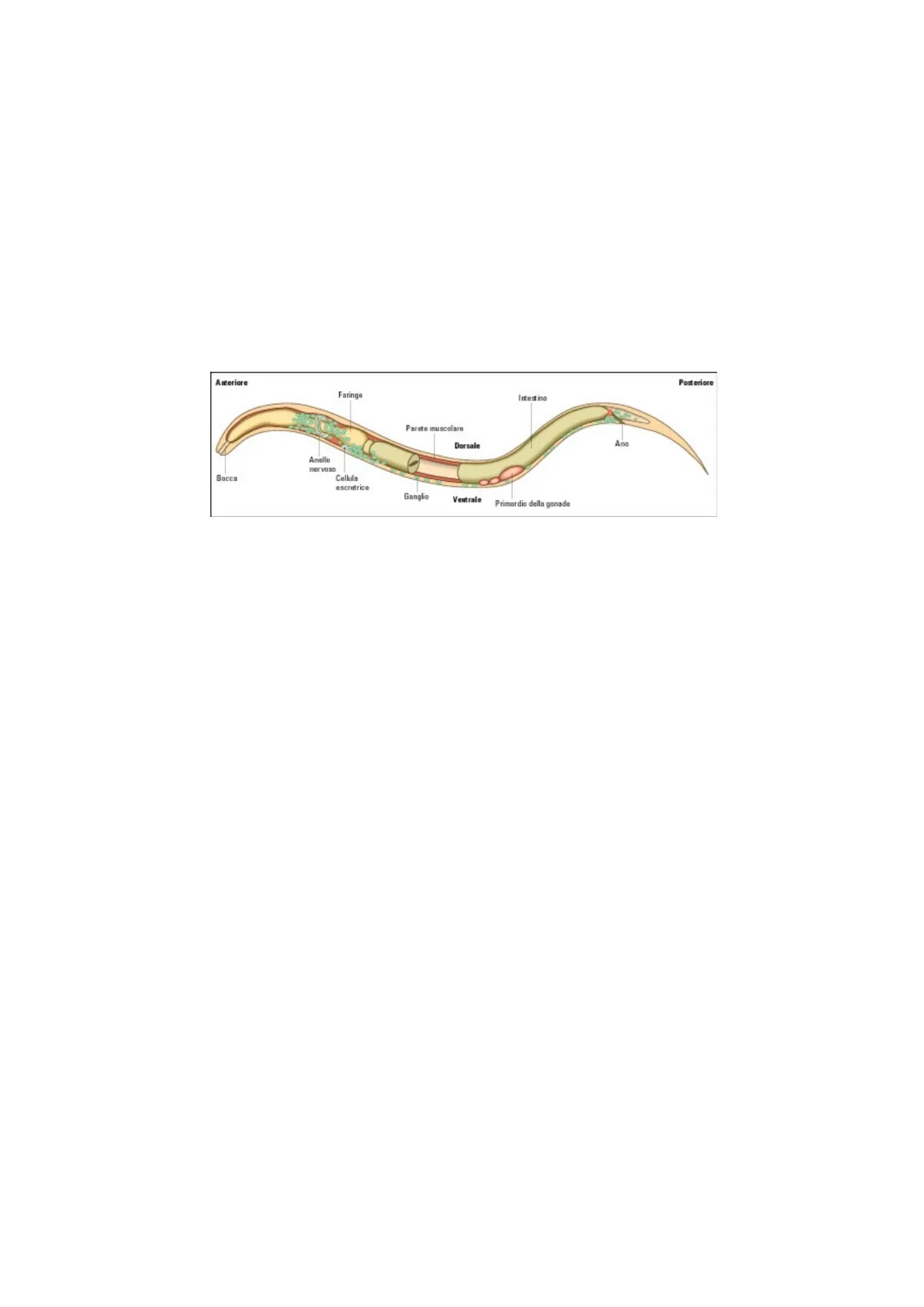
Il motivo per il quale Caenorhabditis Elegans si presta bene allo studio apoptotico è il fatto che tutti gli individui sono
caratterizzati da un numero predefinito di cellule (dallo zigote se ne sviluppano 1090, ne muoiono 131 e ne sopravvivono
959). Conoscendo questo numero predefinito è possibile tracciare ciò che la cellula diventa nell'individuo adulto, compreso
il fatto che alcune di queste muoiono.
Per Elegans sono stati identificati 3 geni mutanti, che poi in seguito sarebbero diventati 4. 2 di questi, il ced-3 ed il ced-4
determinavano la sopravvivenza di cellule. Il terzo gene, il gene ced-9, determinava la morte di tutte le cellule. Ced-3 e ced-
4 sono necessari per far avvenire l'apoptosi, mentre ced-9 la inibisce.
Anteriore
Posteriore
Faringe
Intestino
Parete muscolare
Ano
Dorsale
Anello
nervoso
Bocca
Cellula
escrotrice
Ganglio
Ventrale
Primordio della gonade
Le caspasi
Andando avanti negli studi si è visto che i geni che erano stati identificati in Elegans erano conservati anche nei mammiferi.
In particolare, si è visto che ced-9 era corrispondente alla proteina bcl-2 e che ced-3 era l'equivalente di un gene che
chiameremo Apaf-1 di una famiglia che chiameremo delle caspasi.
Le caspasi sono la famiglia proteica principale che si occupa dell'apoptosi cellulare. Il suffisso -asi ci fa capire che si tratta di
enzimi proteolitici, la c sta per cisteina, ossia l'amminoacido presente nel centro reattivo di queste proteine. Infine, per asp
si intende il fatto che queste riconoscono un residuo di acido aspartico e tagliano le proteine bersaglio a livello di
quest'ultimo.
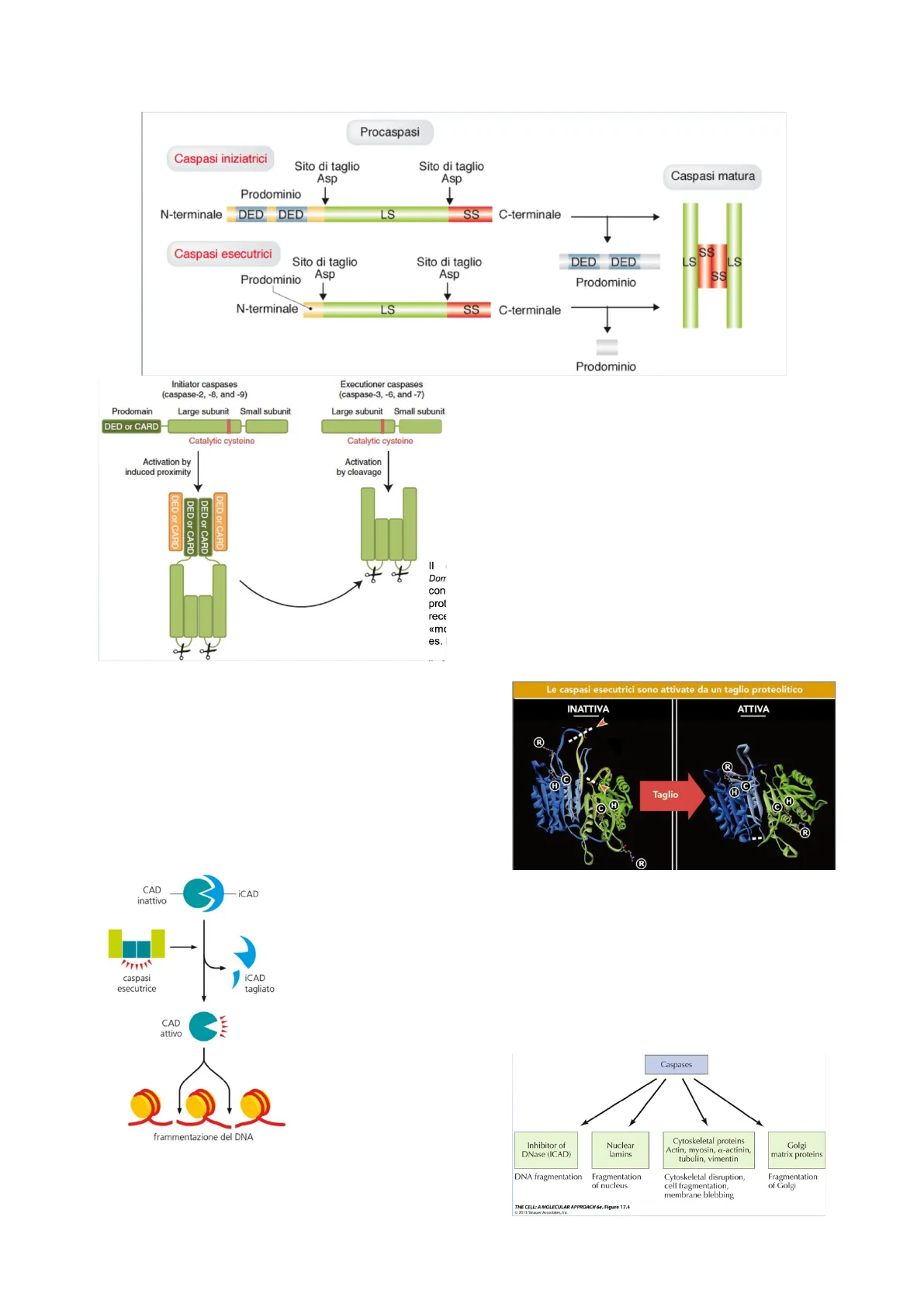
Le caspasi, in quanto proteine di taglio, sono pericolose e vanno quindi gestite e per questo motivo vengono sintetizzate
come procaspasi inattive; quindi il gene produce dei polipeptidi lunghi che sono inattivi e sono caratterizzati da quello che
viene definito un prodominio allele terminale, che è un regolatore negativo che spegne la caspasi; il resto del polipeptide,
perché le caspasi siano attive deve essere processato in due sottounità. Le procaspasi per diventare caspasi hanno quindi
bisogno di due tagli: un taglio che rimuove il prodominio e un altro taglio sulla parte restante della proteina a livello della
subunità. Entrambi i polipeptidi costituiscono la caspasi attiva; infatti poi questi si uniscono a formare un tetramero
costituito da due caspasi processate, definendo la caspasi attiva stessi.Procaspasi
Caspasi iniziatrici
Sito di taglio
Asp
Sito di taglio
Asp
Caspasi matura
Prodominio
-
N-terminale
DED
DED
LS
SS
C-terminale
Caspasi esecutrici
Sito di taglio
Asp
Sito di taglio
Asp
DED
DED
LS
LS
Prodominio
Prodominio
N-terminale
LS
SS
C-terminale
Initiator caspases
(caspase-2, -8, and -9)
Executioner caspases
(caspase-3, -6, and -7)
Prodomain
Large subunit Small subunit
Large subunit Small subunit
DED or CARD
Catalytic cysteine
Catalytic cysteine
Activation by
induced proximity
Activation
by cleavage
DED or CARD
DED or CARD
DED or CARD
DED or CARD
of
=
Dom
con
prot
rece
«mc
es.
of of
Le caspasi sono suddivisibili in due gruppi principali, ossia le
caspasi iniziatrici e le caspasi esecutrici. Le prime
intervengono all'inizio del processo caspasico, mentre le
esecutrici alla fine. Inoltre, ci sono delle caspasi che non sono
richieste
nei
processi apoptotici ma
agiscono
nell'infiammazione, le quali prendono proprio il nome di
caspasi infiammatorie.
Ritornando sulle caspasi iniziatrici, c'è da dire che queste nel
prodominio hanno delle sequenze amminoacidiche
conservate (il DED domain e il CARD domain) che servono
per la loro attivazione. Delle caspasi esecutrici è doveroso
sottolineare che invece non hanno questi domini.
I domini DED e CARD servono per l'attivazione delle caspasi
iniziatrici che li hanno, poiché la loro attivazione dipende
dall'interazione che le procaspasi iniziatrici hanno con delle
proteine adattatrici che si trovano o in membrana o nel citoplasma.
Le caspasi esecutrici, invece, vengono attivate direttamente dal
taglio proteolitico delle iniziatrici.
CAD
inattivo
-iCAD
caspasi
esecutrice
iCAD
tagliato
CAD
attivo
frammentazione del DNA
nucleo
su
sé
stesso
frammentandosi. Un ulteriore bersaglio sono le proteine del
citoscheletro e le proteine della matrice del Golgi.
Le caspasi esecutrici sono attivate da un taglio proteolitico
INATTIVA
ATTIVA
R
R
H
C
H
Taglio
C
®
Le caspasi esecutrici vanno a tagliare le proteine ICAD, che sono degli inibitori di
proteine che sono delle nucleasi del DNA.
È possibile vedere che una proteina CAD taglia il DNA e che questa in una cellula vitale
è inibita da un inibitore che si chiama
iCAD. Nel momento in cui le caspasi sono attive, il loro bersaglio è iCAD, che quindi
viene rimosso e la DNasi può tagliare il DNA.
Altro bersaglio delle caspasi
Caspases
sono le lamine nucleari; le
caspasi smembrano la lamina
nucleare facendo collassare il
Inhibitor of
DNase (ICAD)
Nuclear
lamins
Cytoskeletal proteins
Actin, myosin, a-actinin,
tubulin, vimentin
Golgi
matrix proteins
DNA fragmentation
Fragmentation
of nucleus
Cytoskeletal disruption,
cell fragmentation,
membrane blebbing
Fragmentation
of Golgi
THE CELL: A MOLECULAR APPROACH 6e, Figure 17.4
O 2013 Sinauer Associates, Inc.
Prodominio
SS
SS
Vie di attivazione dell'apoptosi - via estrinseca
L'apoptosi può avvenire per via di un segnale esterno alla cellula (via estrinseca dell'apoptosi) oppure per via di un segnale
interno (via intrinseca dell'apoptosi). Vediamo adesso la via estrinseca dell'apoptosi.
All'esterno della cellula sono presenti recettori di morte (FAS, TNF-R1, TRIAL-R1/2), costituiti da proteine transmembrana,
caratterizzati da una regione extra-cellulare capace di legare i ligandi. Questi recettori di morte hanno un unico dominio
transmembrana di 60/80 amminoacidi, che prende il nome di dominio di morte.
TRAIL e recettori
TRAIL
1
TRAIL-R1/2
Il segnale di morte, detto TRIAL, si lega al recettore che si trimerizza. A questo
punto nella porzione citoplasmatica avvengono dei cambiamenti conformazionali
che permettono a questo recettore di reclutare una proteina adattatrice
chiamata FADD. Quest'ultima da una parte lega il recettore, mentre dall'altra
recluta le proteine caspasi iniziatrici (quest'ultime non possono legarsi
direttamente al recettore, quindi si legano al FADD che fa da ponte). Nel
momento in cui due proteine iniziatrici si associano al FADD, ciascuna taglia il
prodominio dell'altra. Una volta che questo è stato tagliato, queste si tagliano
ulteriormente, producendo quindi una caspasi attiva iniziatrice, la quale andrà a
tradurre gli effettori. Tendenzialmente la procaspasi inizatrice della via estrinseca
è la procaspasi-8, che poi andrà a tagliare la caspasi-3.
Ci sono dei virus che una volta entrati nella cellula non hanno intenzione di farla morire e quindi producono delle proteine
che fanno ad impedire l'associazione tra FADD e le caspasi iniziatrici, proteggendo la cellula dai processi apoptotici.
Segnale di morte TNF
Un altro segnale di morte è il TNF. Per questo segnale sono presenti due
tipi di recettori: il recettore di tipo 1 e il recettore di tipo 2. Questi ultimi
sono espressi nelle cellule del sistema immunitario e rispondono al TNF
solo nella sua forma solubile e diffusibile. I recettori di tipo 1 che sono
espressi nella gran parte dei tessuti rispondono sia alla forma solubile
del TNDF che alla sua forma di membrana.
Il recettore di tipo 2 funziona nel seguente modo: si associa al recettore
attivando la chinasi Ikb, che disattiva l'inibitore di NF-KB. Quest'ultimo
poi può traslocare sul DNA e svolgere il suo ruolo di regolatore della
trascrizione.
TNF&
TNF&
receptor -
IKB
NEMO
NFKB
activated
IKK
complex
IKK COMPLEX
PHOSPHORYLATES IKB
P
P
UBIQUITYLATION
AND DEGRADATION< />OF PHOSPHORYLATED
IKB IN PROTEASOMES
coactivator
protein
TRANSLOCATION OF
NFKB INTO NUCLEUS
-
TRANSCRIPTION OF
NFKB TARGET GENES
Trascrizione di IkB che
spegne il segnale del TNF
DD
DD
FAD
DID
DED
Caspase 8/10
DED H DED
DED
DED DED
DISC
(Death-
Inducing
Signaling
Complex).
APOPTOSIS
DED DED H
active Caspase 8/10
LIBERATION
OF NFKB
Figure 15-62 Molecular Biology of the Cell 6e ( Garland Science 2015)
Recettore di tipo 1
TNF
-TNFR1
Membrana
plasmatica
Domini di morte
FADD
Procaspasi-8
TRADD
Caspasi-8 iniziatrice
Procaspasi esecutrice
Caspasi esecutrice
APOPTOSI
Il recettore di tipo 1, invece, si lega al TNF e trimerizza e scarica una proteina
inibitoria chiamata SODD, formando un complesso di due proteine, FADD e
TRADD. La prima recluta e attiva una procaspasi-8, che induce a sua volta la
propria attivazione autoproteolitica e il susseguente clivaggio di altre caspasi
effettrici che inducono il processo apoptotico.
Recettore FAS o CD95
Altro recettore di morte è FAS o CD95. Le cellule che
esprimono questo recettore vengono rimosse da cellule
che esprimono FASL (ligando di FAS). Nell'associazione
ligando-recettore, quello che succede è che muore la
cellula che esprime FAS.
-Fas ligand
-Fas
Cytosol
-Adaptor
Caspase-8
Autocleavage
Active
caspase-8
Downstream
caspases
killer lymphocyte
Fas ligand
target cell Fas death receptor
TTT
FADD death domain
adaptor
protein
DISC
activated
caspase-8 or 10
death effector
domain
procaspase-8 or 10
activation of
executioner
caspases
apoptotic
target cell
ASSEMBLY
OF DISC
ACTIVATION AND
CLEAVAGE OF
PROCASPASE-8,-10,
OR BOTH
DISC: Death Inducing Silencing
Complex
Questo succede per esempio con i linfociti T
Cell death
citotossici, i quali esprimono in membrana i
ligandi di FAS. Nel momento in cui identificano una cellula che
esprime FAS o CD95, procedono all'associazione tra il linfocita T e la
cellula CD95. L'associazione procede nello stesso modo che
abbiamo visto prima: il recettore trimerizza, chiama FADD che a sua
volta chiama le procaspasi iniziatrici, che a loro volta attivano le
esecutrici. Il complesso che si va a a formare viene detto DISC
complex.
Vie di attivazione dell'apoptosi - via intrinseca
Analizziamo adesso la via di attivazione intrinseca dell'apoptosi. Questa dipende dai mitocondri, in particolare dal rilascio
di citocromo C nel citoplasma della cellula. Ritornando al modello di Elegans, abbiamo determinato questo modello
comprendendo cosa fanno le caspasi. La via intrinseca ci informa su quelli che sono gli altri geni che sono conservati in
Elegans e nei mammiferi, in particolare la famiglia delle proteine Bcl-2, le quali regolano la via intrinseca dell'apoptosi e che
è una famiglia proteica in cui le proteine si dividono in tre gruppi funzionali a seconda del numero di domini di interazione
proteina- proteina, i quali porendono il nome di domini SH. Le proteine bcl-2 che presentano quattro domini SH sono
proteine anti-apoptotiche, quelle che ne presentano tre sono pro-apoptotiche e infine quelle che ne presentano solo uno
(le proteine BH3-only) che sono l'"ago della bilancia".
-death effector
domain
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
