Istologia ed Embriologia: Contrazione Muscolare e Tessuto Nervoso
Documento di Appunti sull'Istologia ed Embriologia – Lezione 22. Il Pdf, un documento di Biologia per l'Università, esplora la contrazione muscolare e introduce il tessuto nervoso, analizzando gli eventi elettrici e fisico-chimici e i meccanismi di attivazione del muscolo liscio.
Mostra di più25 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Contrazione Muscolare e Introduzione al Tessuto Nervoso
Istologia ed Embriologia - Lezione 22 Sbobinatore: Infanti Mauro Controsbobinatore: Preluca Cosmin Docente: Giuseppe Bonaventura
La successione degli eventi sia elettrici che fisico-chimici che caratterizzano la contrazione muscolare.
lucleo Tubulo trasverso Cisterna terminale del reticolo sarcoplasmatico Sarcolemma Terminazione nervosa motoria Miofibrilla 0000010 Mitocondrio
Nell'immagine potete vedere una fibra muscolare tagliata in sezione. Affinché questa fibra muscolare possa attivarsi e quindi contrarsi, deve ricevere sul sarcolemma, cioè sulla sua membrana plasmatica un input, vale a dire una sollecitazione o un segnale. Questo segnale giunge per mezzo di una terminazione nervosa di tipo motorio (che parte da un motoneurone), la quale si adagia sul sarcolemma, ponendosi quindi a contatto con la membrana dove va a formare una giunzione sinaptoide. Questa terminazione riesce a provocare l'insorgenza sul sarcolemma di un potenziale d'azione, vale a dire un'onda di depolarizzazione che causa lo sconvolgimento della distribuzione delle cariche elettriche nei due versanti della membrana.
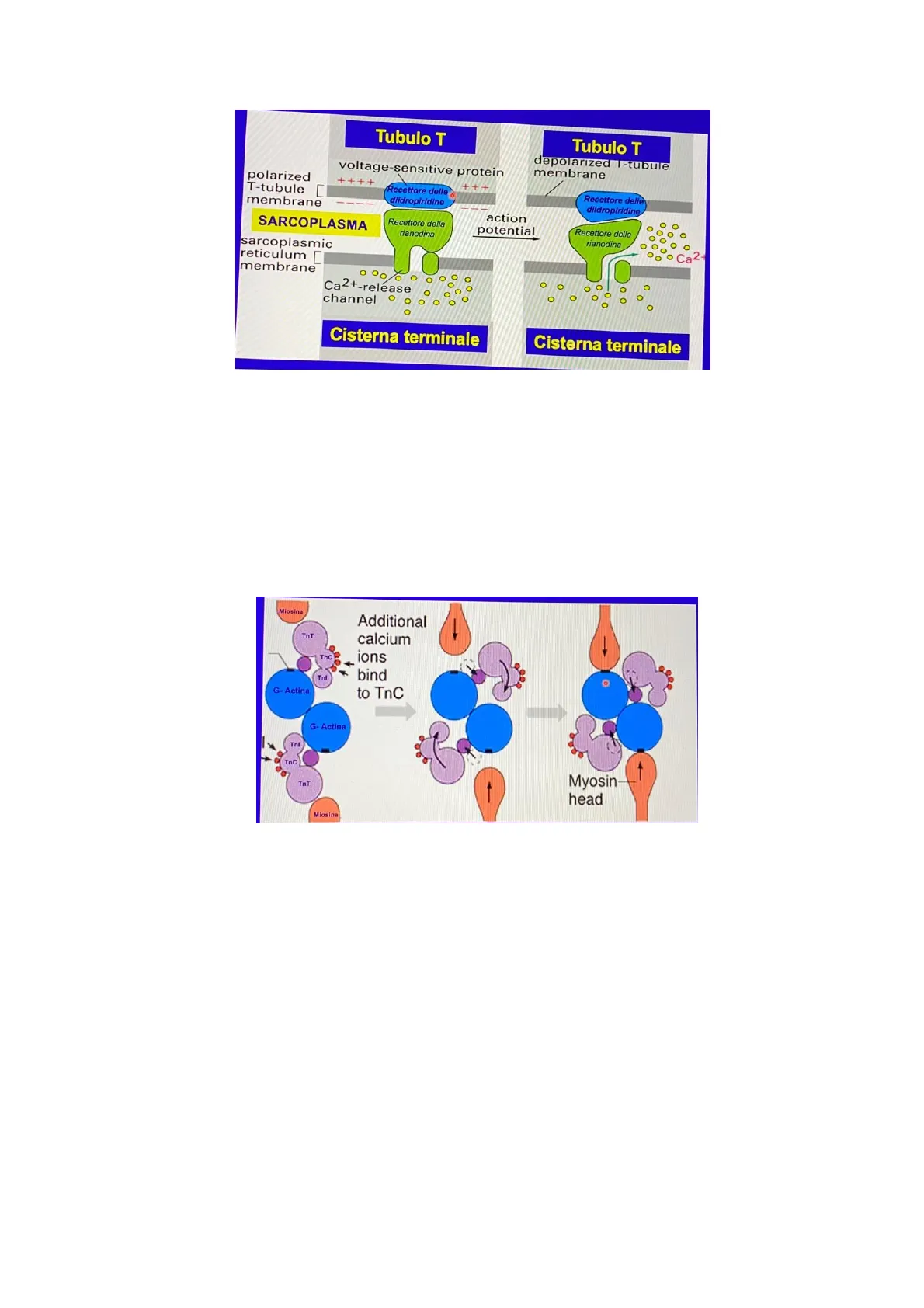
Una volta che quest'onda di depolarizzazione si è avviata, essa comincia a viaggiare e a propagarsi lungo tutto il sarcolemma e anche nel sottostante sarcoplasma per mezzo dei tubuli T (invaginazioni del sarcolemma). Questo segnale, dunque, si porta anche a ridosso delle cisterne terminali (i tubuli T si sviluppano e si posizionano a fianco di due cisterne terminali di due sistemi sarcotubulari adiacenti).Tubulo T voltage-sensitive protein Tubulo T depolarized I-tubule membrane polarized T-tubule membrane ++++ +++ Recettore delle diidropiridine Recettore delle diidropiridine SARCOPLASMA Recettore della rianodina action potential sarcoplasmic reticulum membrane Recettore della rianodina Ca2 Ca2+-release channel Cisterna terminale Cisterna terminale
Quando quest'onda di depolarizzazione si propaga lungo la membrana del tubulo T (quello azzurro in figura), si viene a determinare una modifica conformazionale della proteina voltaggio-sensibile noto come recettore delle diidropiridine, sulla membrana del tubulo T. Per via di questo cambio conformazionale, essa provoca a sua volta l'apertura del canale del calcio, il recettore della rianodina, posizionato sull'adiacente membrana della cisterna terminale del reticolo sarcoplasmatico. L'apertura del canale del Calcio permette la fuoriuscita dalle cisterne di grandi quantità di ioni calcio, i quali potranno quindi diffondersi liberamente nel sarcoplasma.
Disponibilità di Calcio e Interazione Miosina-Actina
Miosina TnT TnC Tnl G- Actina G- Actina Tnl Tnc t TnT t Myosin head Miosina Additional calcium ions bind to TnC
L'improvvisa disponibilità di massicce dosi di calcio (pallini rossi nell'immagine) nel sarcoplasma, ha come conseguenza il suo legame alla subunità TnC della troponina, che è fortemente affine al Calcio. Questo legame, a sua volta, si rende responsabile di un cambio di conformazione spaziale dell'intero complesso della troponina. La troponina cambia forma, e siccome ha attaccata la tropomiosina, questo cambio conformazionale sposta la tropomiosina dai siti attivi presenti sulle molecole di G-Actina, rendendo questi siti attivi disponibili. A questo punto, la testa della miosina può avanzare e può interagire con il sito attivo della G-Actina che è rimasto scoperto.
Semplificando, si potrebbe dire che il ruolo funzionale di queste due proteine (troponina e tropomiosina) è quello di reagire alle variazioni della concentrazione di calcio.
Possiamo immaginare queste due proteine come una sorta di interruttore, on-off: se c'è calcio si attivano e può avvenire la contrazione muscolare, se non c'è calcio l'interruttore è spento e la contrazione muscolare non può avvenire.
Rappresentazione Schematica della Contrazione Muscolare
Fase di Riposo della Miosina
A riposo, la testa della miosina è inclinata a 45° rispetto al filamento sottile e porta sempre legata una molecola di ATP ATP 0 a
La molecola di miosina porta sempre legata una molecola di ATP.
La testa della miosina, quando è a riposo, è staccata dal filamento sottile ed è inclinata di 45 gradi rispetto al filamento sottile.
Idrolisi dell'ATP e Rotazione della Testa Miosinica
L'arrivo del Calcio, facendo venire meno l'inibizione esercitata dalla Tnl sulla ATPasi del frammento S1, provoca l'idrolisi dell'ATP, ma sia l'ADP che il fosfato rimangono attaccati sul frammento S1; l'energia liberatasi fa ruotare la testa della miosina (90°) ADP+Pi b
Con la disponibilità dello ione calcio, che è fuoriuscito dal reticolo, la testa miosinica può idrolizzare l'ATP. Questo perché il calcio per prima cosa si v a legare alla TnC modificando la conformazione spaziale della troponina. Per questo motivo viene meno quella inibizione che la subunità Tnl esercita sull'adenosintrifosfatasi della testa miosinica (la testa miosinica è dotata di attività adenosintrifosfatasica che è inibita dalla subunità TnI, quella inibitrice). Cambiando conformazione, la troponina non solo modifica la posizione della tropomiosina ma viene anche meno questa funzione inibitrice svolta dalla Tnl nei confronti dell'attività adenosintrifosfatasica della testa miosinica. A questo punto l'adenosintrifosfatasi può agire e può portare all'idrolisi dell'ATP, che comporterà la formazione di ADP + fosfato inorganico (Pi), inizialmente attaccati alla testa miosinica. L'idrolisi dell'ATP ha comportato una liberazione di energia e questa energia, la testa della miosina l'ha sfruttata per ruotare. Questa energia provoca una rotazione della testa miosinica che si è posta a 90 gradi rispetto al miofilamento sottile.
Legame della Testa Miosinica all'Actina
La testa della miosina (90°) si lega al filamento di actina ADP+Pi C
Con questa inclinazione di 90 gradi, la testa miosinica può finalmente andarsi a legare ad uno dei siti attivi della G-Actina presenti sul miofilamento sottile.
Rilascio di ADP e Pi e Colpo di Frusta
ADP+Pi d L'interazione acto-miosinica favorisce il rilascio di ADP e Pi, con conseguente brusco ritorno della testa miosinica alla configurazione a 45° (colpo di frusta), che determina il trascinamento del filamento sottile verso il centro del sarcomero.
Questo legame miosina-actina favorisce il rilascio di ADP e Pi, cio è responsabile di un nuovo cambiamento conformazionale della testa miosinica, la quale ritorna alla configurazione che aveva prima, cioè a 45 gradi. Durante questa flessione, la miosina subisce il "colpo di frusta", cioè un movimento improvviso che sposta il filamento di actina, trascinandolo verso il centro del sarcomero.
Quindi il filamento sottile si sposta perché la testa della miosina aggancia uno dei siti attivi della G- Actina, poi ruota, flette di 45 gradi e quindi trascina il filamento di actina.
Scivolamento dei Filamenti Sottili
thin filament thick filament 1 Le linee Z si avvicinano trascinate dai filamenti sottili che scivolano sui filamenti spessi Z WISC
La motivazione per cui i dischi Z di un sarcomero si avvicinano tra di loro, è proprio perché sono trascinati dallo scivolamento telescopico dei filamenti sottili sui filamenti spessi.
Ciclo di Contrazione e Rigidità Cadaverica
ATP D a
Alla fine della contrazione si ritorna alla situazione iniziale e il ciclo si rinnova e questo avviene nel momento in cui si ha l'aggancio di una nuova molecola di ATP al frammento S1, cosa che contestualmente provoca il distacco, la dissociazione tra actina e miosina. La testa della miosina, dunque, risulta nuovamente disponibile per interagire con un nuovo sito di una molecola G-Actina.
È proprio la disponibilità di ATP a permettere questa dissociazione tra actina e miosina, tanto è vero che quando un individuo muore, in un tempo variabile compreso tra 5-7-12 ore, si ha l'esaurimento delle scorte di ATP e actina e miosina non si dissociano più, anzi si formano dei complessi stabili, per cui tutto questo è responsabile della "rigidità cadaverica".
È ovvio che, affinché un muscolo possa contrarsi in modo efficace, è necessario che questa successione di eventi si debba ripetersi in maniera rapidissima tantissime volte, il che è possibile fin tanto che c'è disponibilità di ioni calcio.
Cessazione dell'Impulso Nervoso
Una volta che è cessato l'impulso nervoso (quando il nostro cervello non deve più comandare un muscolo di contrarsi) si dovranno verificare fenomeni opposti rispetto a quelli visti durante la contrazione.
- Abbiamo un richiamo di calcio all'interno delle cisterne del reticolo sarcoplasmatico. Questo avviene perché vengono attivate delle pompe del calcio, che sono installate sulla membrana del reticolo sarcoplasmatico. Si attivano queste pompe che iniziano a richiamare calcio e lo riportano all'interno del lume del reticolo sarcoplasmatico.
- La conseguenza è che si ha una diminuzione del calcio nel sarcoplasma e questo porta al distacco dello ione calcio dalla subunità TnC della troponina. La troponina, dunque, riasume la sua conformazione spaziale originaria e questo riporta la tropomiosina al proprio posto.
- La tropomiosina torna nella sua posizione originaria mascherando i siti attivi della G-actina per la miosina.
- Cessazione della contrazione
Giunzione Neuromuscolare
.GIUNZIONE NEUROMUSCOLARE FN Fibra muscolare GN
Questa è un'immagine, ottenuta al microscopio elettronico a scansione, di una giunzione neuromuscolare o placca motrice. GN sta per "giunzione neuromuscolare". FN sta per "fibra nervosa", è una fibra nervosa efferente di tipo motorio.
Struttura della Fibra Nervosa Motrice
Ramificazioni nervose terminali Nucleo della cellula di Schwann Assone Vescicole sinaptiche Pieghe giunzionali guaina mielinica Miofibrille Nucleo muscolare
La fibra nervosa motrice, che è formata da un assone o cilindrasse rivestito da guaina mielinica, in prossimità della sua estremità terminale perde la guaina mielinica (infatti si dice che l'assone resta nudo) e non solo, accade anche che l'estremità terminale dell'assone si arborizza, ovvero si divide in piccole ramificazioni, le ramificazioni nervose terminali. Sono i così detti "rami terminali"
Rami Terminali e Bottone Sinaptico
Ramificazioni nervose terminali Nucleo della cellula di Schwann Assone Vescicole sinaptiche Pieghe giunzionali Rami terminali Miofibrille Nucleo muscolare
L'assone che si è ramificato in realtà non è completamente nudo perché tutti questi rami terminali risultano rivestiti dal citoplasma della cellula di Schwann (è una cellula gliale, della neuroglia, che nel Sistema Nervoso Periferico si occupa di produrre il rivestimento mielinico). Manca la guaina mielinica ma non c'è una nudità assoluta.
Ognuno di questi rami terminali termina con un'estremità un po' più globosa, che è detta "bottone sinaptico". Questo bottone sinaptico è accolto da un affossamento che si forma sulla superficie della fibra muscolare.
Doccia o Fessura Sinaptica
Doccia o fessura o spazio sinaptico: Doccia o fessura sinaptica 6 Membrana postsinaptica Membrana presinaptica
La giunzione neuromuscolare o placca motrice è la zona di contiguità tra la membrana del bottone sinaptico (quella in rosso) e il sarcolemma (quello in verde) della fibra muscolare. In questa sede possiamo distinguere tre diverse regioni:
- Membrana presinaptica: la porzione di membrana del terminale assonico che è coinvolta nella giunzione;
- Il sarcolemma che forma la membrana postsinaptica;
- Doccia o Fessura Sinaptica: spazio che si viene a determinare tra queste due membrane.
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
