Proteínas del Citoesqueleto: Actina y Miosina en la Contracción Muscular
Documento de Universidad sobre Proteínas Del Citoesqueleto. El Pdf, un material esquemático y detallado, explora la estructura y función de la actina y miosina, así como los mecanismos de contracción muscular, siendo útil para estudiantes de Biología.
Ver más18 páginas


Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Proteínas del Citoesqueleto
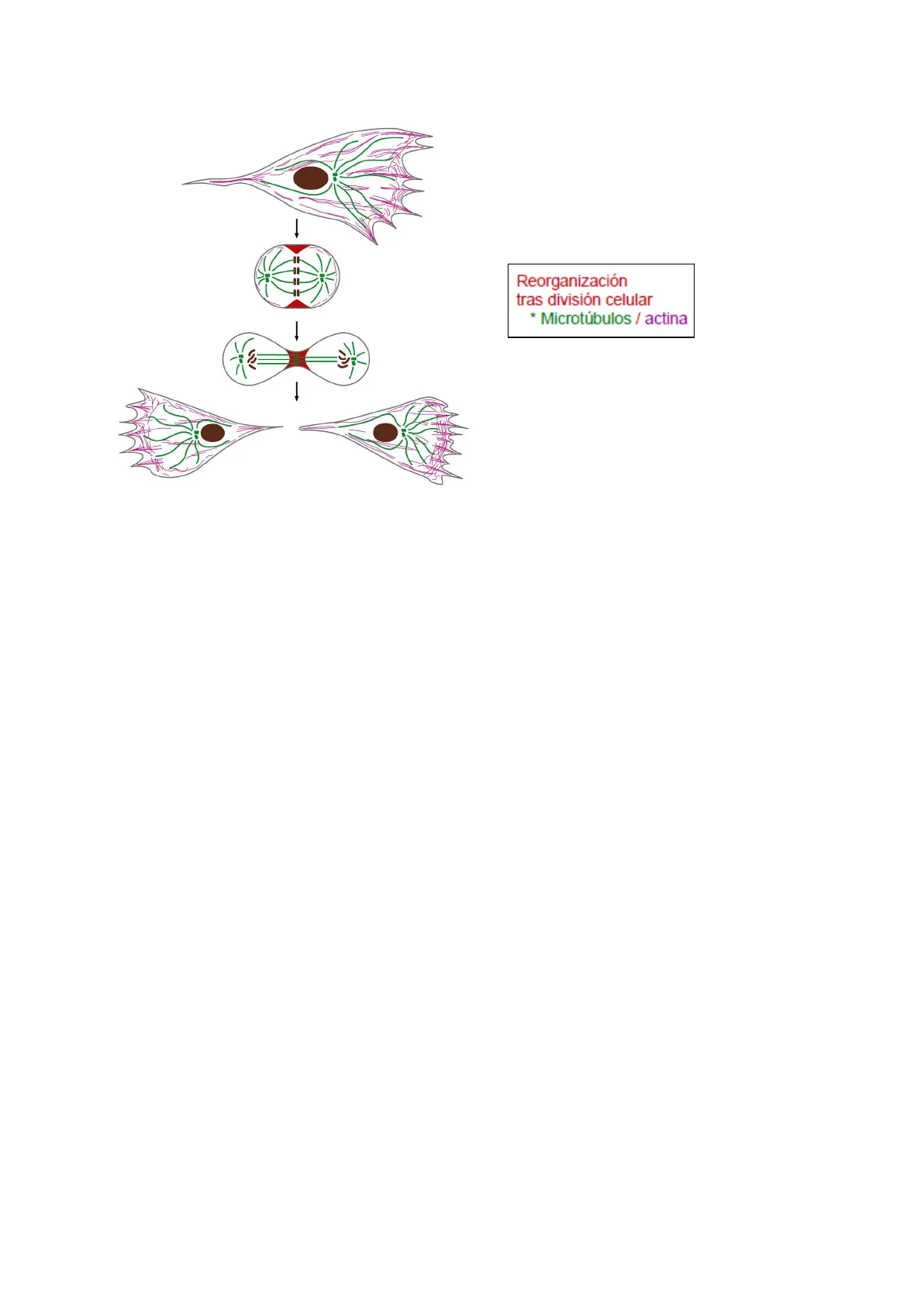
La célula para desarrollar su función necesita:
Forma adecuada de acuerdo a la función que desempeña
- Organización interna del citoplasma celular, organización de orgánulos concreta.
-> Transporte vesicular y desplazamiento de orgánulos: debe desplazar vesículas y
orgánulos, por lo que es dinámica y necesita estar en contacto con otras (de lo contrario,
apoptosis): Interacción con entorno y células vecinas.
> Resistencia: debe resistir esfuerzos y tensiones
➔
Reorganización tras división: ser capaz de mover la célula y reorganizar las células hijas
tras la división.
Todas estas funciones las realizan un gran número de proteínas que constituyen el citoesqueleto
celular.
Citoesqueleto: Red de Elementos Proteicos
Es una compleja red de elementos proteicos del citoplasma de la célula. Está constituido por 3
redes complejas:
- Microfilamentos de actina (8 nm): son los más finos. Tienen membrana apical, basal y
lateral. Rellena las microvellosidades, forma cortex celular, en uniones adherentes,
citocinesis. - Filamentos intermedios (10 nm): tamaño intermedio. Suelen estar entre las
membranas laterales. Unidos a desmosomas - Microtúbulos (25 nm): de tamaño mayor y se superponen en todo el citosol celular.
Forman cilindros huecos de tubulina (en los laterales). Sirven de guía en el transporte
vesicular y de orgánulos y forma el huso mitótico tirando de los cromosomas
25 um
25 um
25 um
Tipos de Filamentos Celulares
FILAMENTOS INTERMEDIOS
Diferentes según el tipo
celular (10 nm)
MICROTÚBULOS
Tubulina (25 nm)
FILAMENTOS DE ACTINA
Actina (8 nm)Reorganización
tras división celular
- Microtúbulos / actina
Funciones del Citoesqueleto Celular
EL CITOESQUELETO CELULAR PARTICIPA EN ... (7)
Forma
-> Movimiento
-> División
> Interacciones celulares
- Organización interna del citoplasma
>> Desplazamiento de orgánulos celulares
> Transporte vesicular.
Actina y Proteínas Asociadas
Los filamentos de actina (también conocidos como microfilamentos) son polímeros helicoidales
de dos hebras de la proteína actina. Tienen la apariencia de estructuras flexibles, con un
diámetro de 5 a 9 nm, y están organizados en una variedad de haces lineales, redes
bidimensionales y geles tridimensionales. Aunque los filamentos de actina están dispersos por
toda la célula, su concentración máxima se encuentra en la corteza, justo debajo de la
membrana plasmática.
Presentes en todos los tipos de células eucariotas, pero es más abundante en células
musculares (10% peso del total de proteínas) que en células no musculares (1 - 5 %).
La actina se une a una gran variedad de proteínas accesorias para participar en diferentes
procesos celulares (mitosis, división celular, polaridad, secreción, endocitosis, síntesis de lípidos,
filamentos dinámicos, .. )ADP
ATP
Polimerización de la Actina
Actina unida
al ADP
Actina unida
al ATP
Pi
POlIMERIZACIÓN DE LA ACTINA
Actina G: monómero globular que contiene un átomo de Mg2+ unido a ATP. Se polimerizan
mediante el enrollamiento de hebras alfa-hélice, para formar:
- F-actina: 2 hebras alfa-hélice, mediante hidrólisis ATP (proceso reversible).
- Subunidades actina: enzimas capaces de hidrolizar ATP a ADP inmediatamente
después de que se une el nuevo monómero. - Cuando unida a ADP, estabilidad reducida del filamento: despolimerización.
Recambio rotatorio.
Al polimerizar los filamentos presentan polaridad en extremos (+) y (-), ya que los monómeros
pueden unirse y separarse por los 2 extremos (principalmente por el extremo + que es el más
dinámico). Se ensamblan por extremo +, se polimeriza la actina / desensamblaje por extremo -
para volver a la estructura actina G.
Sólo permanecen de décimas de segundo a minutos: Recambio rotatorio.
Hay equilibrio dinámico de proteínas G y F en citosol (aproximadamente 50%), excepto en
momentos puntuales. Los monomeros G van formando protofilamentos, convirtiéndose en F, y 2
protofilamentos se enrollan entre sí formando el microtúbulos de actina
extremo más
molécula de actina
NH2
extremo más
COOH
37 nm
ATP
(ADP
cuando
está en el
filamento)
extremo menos
extremo menos
(B
(C)
50 nm
(A)
Tipos de Actina y su Localización
TIPOS DE ACTINA
Altamente conservada. Varios genes codificadores (6 en especie humana): a, ß, y-actina.
- a-actinas: 3 tipos (fibra muscular esquelética, lisa, cardíaca).
- ß -actina y y-actina (no musculares): Forman córtex celular (resistencia mecánica). Esta
red en el polo apical de las células epiteliales aparece terminando en uniones
adherentes y en el polo basal en los contactos focales.
Enlaza con proteínas integrales de membrana a través de proteínas adaptadoras:
Membrana plasmática: espectrina, anquirina y miosina tipo I
Uniones adherentes: vinculina, a-actinina, a-ß-y-cateninas - Contactos focales: a-actinina, vinculina, talina
La actina está asociada a muchas proteínas asociadas para participar en diferentes procesos
celulares.
(D)
(A)
(B)
(C)
Nucleación de Actina Catalizada por el Complejo ARP
ACTINA. NUCLEACIÓN CATALIZADA POR EL COMPLEJO ARP
La activación del complejo ARP requiere la presencia de un factor activador o molécula señal.
Unión ARP - factor activador. Provoca la activación y promueve la modificación de configuración
3D que facilita el crecimiento de los microfilamentos de actina por la adición de monómeros de
actina G en el extremo (+) iniciando la polimerización de la actina F.
La célula recibe una señal de que tiene que nuclear (formar filamentos de actina) por ello el
complejo ARP inactivo se une al factor activador y cambia de conformación. Debido a esto, el
complejo ARP está activo y, por lo tanto, a partir de monómeros de actina (G) estimula la
formación/nucleación del filamento de actina (estos monómeros G polimerizan
formando
monómeros F
dando lugar al filamento de actina).
Se ha visto con los estudios que se han realizado que el
complejo ARP nuclea el crecimiento de un nuevo filamento de
actina de una forma más eficiente cuando está unido
lateralmente a un fragmento preexistente de filamento de
actina F dando lugar a ramificaciones/redes (textura gel).
Este complejo va a ser más activo nucleando cuando justo
está unido a otro filamento ya preformado en un ángulo de 70°.
Esto permite formar redes (esenciales en la formación de
lamelipodios).
70°factor
activador
otras proteínas
extremo
menos
extremo
más
+
Arp3
Arp2
complejo
ARP inactivo
complejo ARP activo
monómeros
de actina
filamento de actina nucleado
actina
Arp2
Arp3
ARP: Proteínas Relacionadas con la Actina
Nucleación de Actina Catalizada por Forminas
ACTINA. NUCLEACIÓN CATALIZADA POR FORMINAS (HACES PARALELOS).
Los dímeros de formina (anillos en verde) se unen en el extremo positivo (donde nuclea el
filamento de actina) y regulan la adición de monómeros de actina G dando lugar a la formación
de haces paralelos (a diferencia del complejo ARP que forma redes).
extremo más
dímero
de formina
filamento
de actina
extremo menos
Fármacos que Afectan el Dinamismo de los Filamentos de Actina
ACTINA. FÁRMACOS QUE AFECTAN AL DINAMISMO DE LOS FILAMENTOS DE ACTINA
COMPUESTOS ESPECÍFICOS DE ACTINA
Faloidina
se une y estabiliza los filamentos
Citocalasina
encasqueta los extremos más de los filamentos
Suinholida
fragmenta filamentos
Latrunculina
se une a las subunidades e impide su polimerización
El conocimiento de procesos naturales nos permite tratar de enfrentarnos a determinadas
patologías.
El conocimiento de la Faloidina (toxina que de forma natural produce la Amanita phalloides
hongo muy venenoso que produce la muerte) nos ha permitido descubrir que se une muy
fuertemente a los filamentos de actina e impide la despolimerización. Es decir, la faloidina
bloquea el recambio rotatorio (polimerización y despolimerización) y, debido a la importancia de
dicho recambio rotatorio, la faloidina resulta ser una toxina/veneno mortal.Ej: comer carne cruda
antídoto natural. Cuando ingerimos la toxina faloidina, esta bloquea el
recambio rotatorio. Sin embargo, cuando consumimos carne cruda añadimos filamentos de
actina, por lo tanto, consumiendo carne cruda conseguimos compensar el efecto de esa toxina.
Citocalasina (hongo). Se une al extremo (+) impidiendo la polimerización de la actina, por lo que
también bloquea el recambio rotatorio.
La suinholida y latrunculina son toxinas producidas por esponjas que también actúan a nivel de
bloqueo de la polimerización / despolimerización de la actina.
Conclusión sobre la Actina
CONCLUSIÓN. Nos interesa que la actina puesto que se tiene que unir a muchas proteínas
accesorias a la actina para participar en diferentes procesos celulares), no ha
evolucionado/modificado mucho para que la puedan reconocer fácilmente todas esas proteínas
asociadas.
Proteínas Asociadas a la Actina
Cap
3
Filamina
Bloqueo de extremo (tapa)
Timosina
6
Enlaces cruzados
2
Secuestro
de monómero
1
Formina
ARP
Fimbrina
Villina
a-actinina
Monómeros
Nucleación
de monómero
Polimerización de monómero
Formación de haces
6
1
Espectrina
Vinculina
7
8
Unión de membrana
5
Despolimerización
Cofilina
Corte de filamentos
Gelsolina + Ca2+
- Cap. Siempre están en los extremos (capuchones), protecciones que bloquean y marcan
dónde están los extremos de los filamentos de actina. - Filamina. Dímero con forma de V que permite formar enlaces cruzados (importante en la
formación de lamelipodios). - Fimbrina, vilina y alfa-actinina permiten la formación de haces paralelos.
- Gelsolina (dependiente de calcio). Corta los filamentos de actina.
- Cofilina. Estimula la despolimerización de los monómeros, es decir, que los monómeros
F pasen a monómeros G (desestabilizan/despolimerizan). - Espectrina. Proteína que ancla el filamento de actina a la membrana plasmática.
4
Diferencias entre Fimbrina y Alfa-Actinina
La fimbrina es más pequeña, forma un haz paralelo pero no deja espacio para que entre los
filamentos de actina entre la miosina que es la proteína motora. Sin embargo, la alfa-actinina es
más grande y deja más hueco entre los filamentos de actina permitiendo la entrada de la
miosina (necesaria para los haces contráctiles y que se lleve a cabo la contracción muscular).
Filamina y Metástasis
La filamina permite hacer entrecruzamientos y la formación de redes que conforman los
lamelipodios. El conocimiento de la filamina llevó a que unos investigadores en el laboratorio
hicieran que disminuyera (dado que la filamina está involucrada en la formación de los
lamelipodios y éstos favorecen el desplazamiento celular). Lo que hicieron fue conseguir en el
laboratorio que unas células de melanoma (tumorales con gran capacidad de metastatización)
disminuyeron la expresión hasta prácticamente anularla de la filamina. La filamina que permite la
formación de los lamelipodios mediante los entrecruzamientos no se estaba expresando. Sin
embargo, en el mismo laboratorio a estas células tumorales se les incentivo la expresión de la
filamina y se vió que sí formaban lamelipodios (se daban entrecruzamientos que estimulaban la
formación de lamelipodios).
(A)
(B)
10 um
Células con burbujeo alrededor (no se expresa la filamina) / células que metastatizan
Si continúa la investigación podríamos llegar a que a un paciente con melanoma podamos
alterar la expresión de la filamina pudiendo FRENAR LA METÁSTASIS.
Proteína ERM y Neurofibromatosis
ERM tiene la misma función que la espectrina (anclan los filamentos de actina a la membrana
plasmática).
¿Por qué es importante ?.
Proteína ERM en estado inactivo
célula recibe señal de que tiene que anclar el filamento de
actina a la membrana plasmática
cambio de conformación (ERM activo)
permite que el
filamento de actina se ancla a la membrana plasmática.
PATOLOGÍA: NEUROFIBROMATOSIS. Producción de muchos tumores benignos de piel que
se ha visto que está asociada a la mutación de una de las proteínas de la familia ERM. No
funciona correctamente el anclaje a la membrana porque la proteína ERM no funciona
correctamente y por lo tanto se producen esos tumores benignos.
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
