Istologia dei tessuti epiteliali: caratteristiche e classificazione
Documento di Università su Istologia dei tessuti epiteliali. Il Pdf, utile per lo studio della Biologia a livello universitario, esplora le giunzioni e la classificazione dei tessuti epiteliali, la polarità cellulare e le diverse tipologie come l'epitelio monostratificato squamoso e quello pseudostratificato.
Mostra di più16 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Caratteristiche dei Tessuti Epiteliali
ISTOLOGIA SILVIA DI AGOSTINO LEZIONE 2 - 08/10/2023 SBOBINATORI: Chiara Caroleo; Adriana Carbonara ARGOMENTI TRATTATI: giunzioni e classificazione dei tessuti epiteliali Le caratteristiche che accomunano tutti i tessuti epiteliali sono le seguenti:
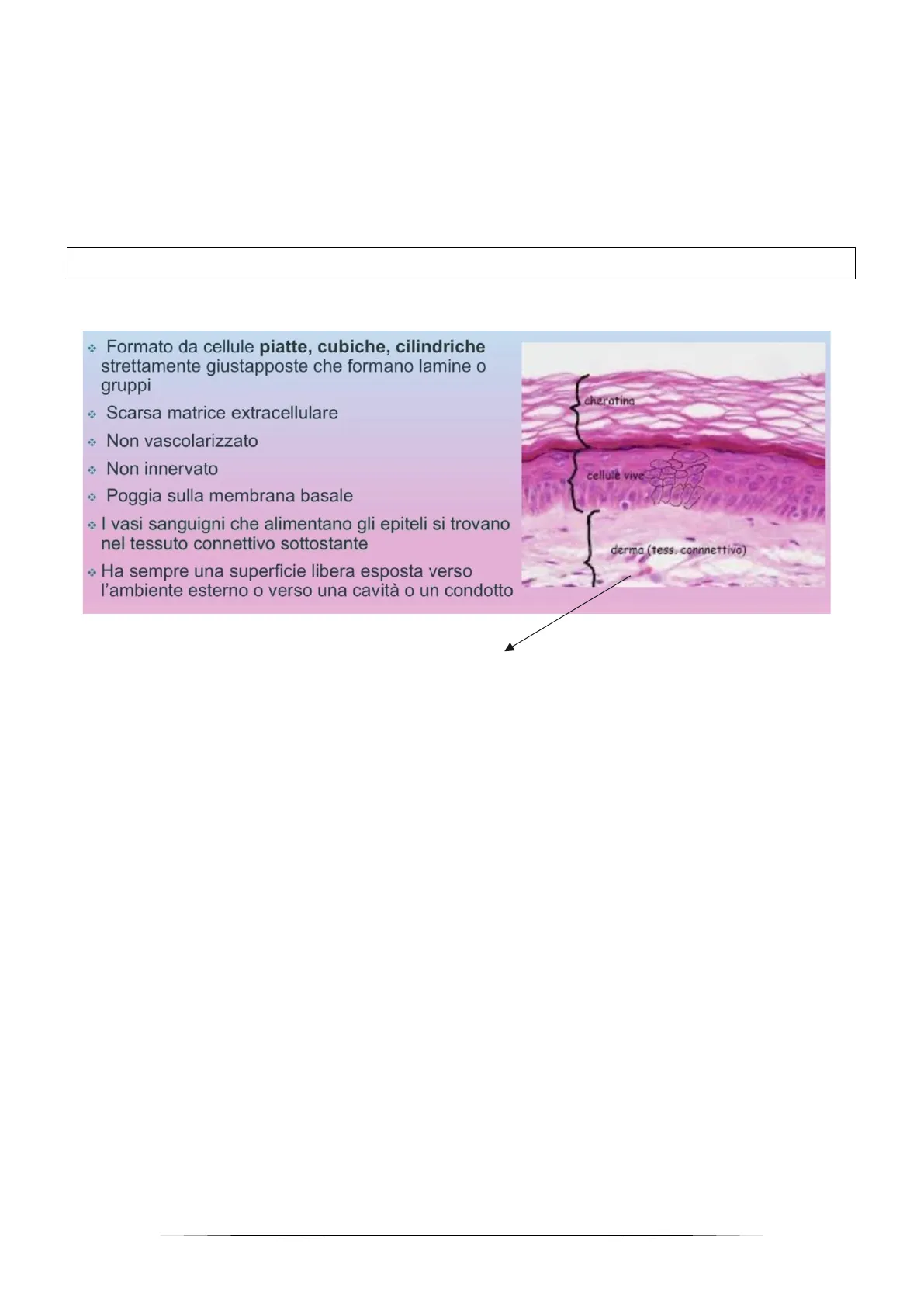
- Formato da cellule piatte, cubiche, cilindriche strettamente giustapposte che formano lamine o gruppi
- Scarsa matrice extracellulare
- Non vascolarizzato
- Non innervato
- Poggia sulla membrana basale
- I vasi sanguigni che alimentano gli epiteli si trovano nel tessuto connettivo sottostante cheratina cellule vive derma (tess. connnettivo)
- Ha sempre una superficie libera esposta verso l'ambiente esterno o verso una cavità o un condotto
Questo preparato istologico è colorato attraverso il metodo ematossilina-eosina, grazie al quale possiamo individuare i nuclei cellulari (colorati di lilla scuro). Nell'immagine sono cerchiate le cellule dell'epidermide, esse, come tutte le cellule epiteliali, appaiono addossate tra di loro e in questo caso formano più strati. Notiamo invece che nel derma, che è un tessuto connettivo, i nuclei sono molto più distanti tra di loro e sono circondati da matrice extracellulare formata da fibre proteiche di varia natura (appare di un rosa molto chiaro poiché l'ematossilina-eosina non è in grado di evidenziare le proteine, a tale fine bisognerebbe utilizzare la colorazione tricromica). Nei tessuti epiteliali tale matrice non è presente, così come le innervazioni e i vasi sanguigni, perciò tutti gli scambi nutritivi avvengono passivamente per diffusione dal tessuto connettivale sottostante. Ciò non toglie che le cellule epiteliali devono comunicare tra di loro e lo fanno grazie alle giunzioni. Osservando l'epidermide possiamo notare che c'è una sorta di polarizzazione tra gli strati. Quando abbiamo a che fare con un multistrato, bisogna distinguere tra lo strato apicale (quello esposto verso l'esterno o verso una cavità) e lo strato basale (cioè quello inferiore che interagisce con la membrana basale). Ad esempio nel caso di una ghiandola lo strato basale è formato dalle cellule più distanti dal lume della ghiandola. Nel caso dell'epidermide, lo strato basale è la sede della nicchia delle cellule staminali, infatti mano a mano che si differenziano si spostano verso l'esterno fino a morire e formare i cheratinociti. Un'altra caratteristica comune a tutte le cellule epiteliali dello strato basale, o comunque dell'unico strato nel caso di un epitelio monostratificato, è che non poggiano direttamente sul tessuto connettivo, ma su una rete di proteine che servono a dare sostegno.
Membrana Basale e Patologie
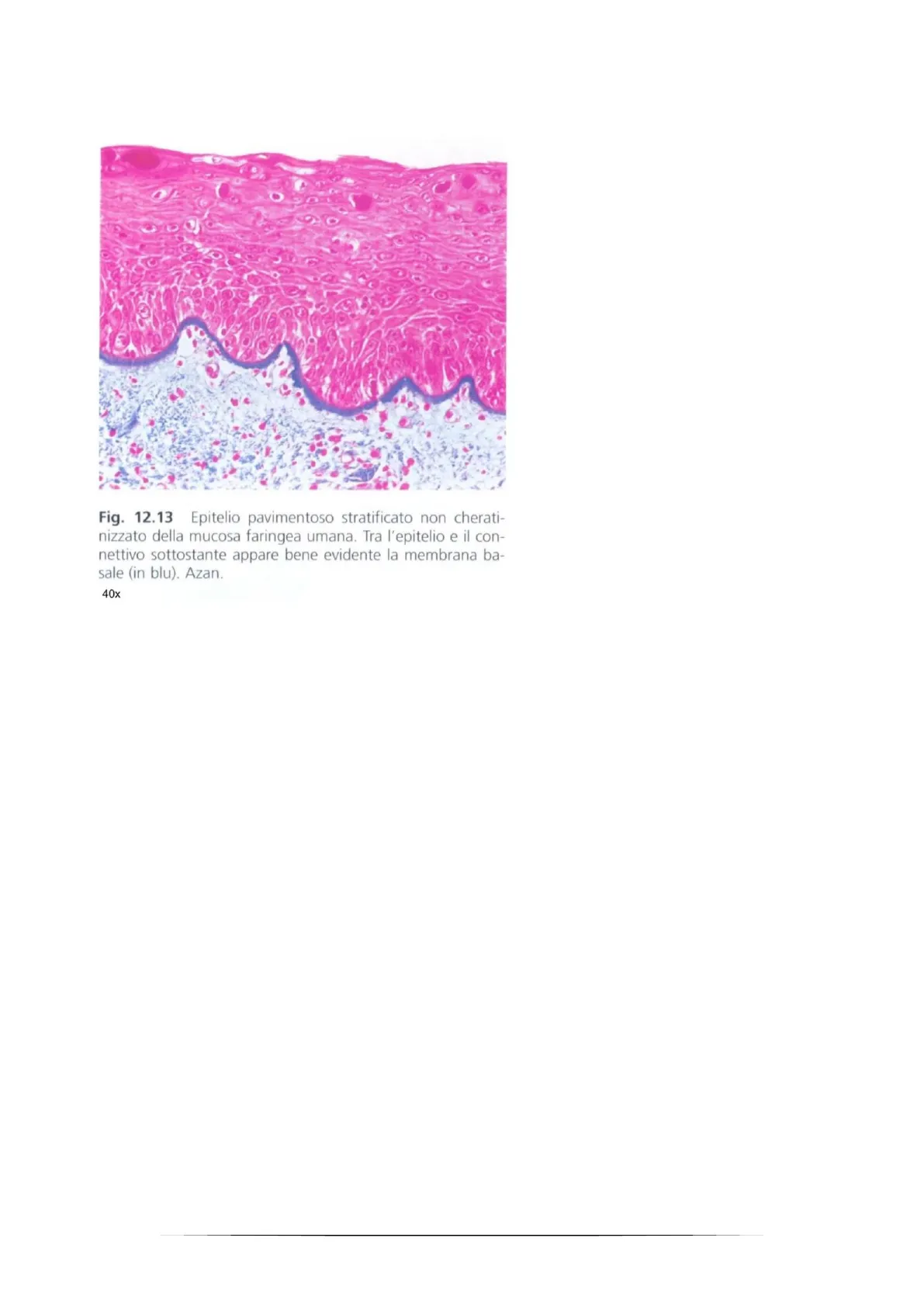
1Fig. 12.13 Epitelio pavimentoso stratificato non cherati- nizzato della mucosa faringea umana. Tra l'epitelio e il con- nettivo sottostante appare bene evidente la membrana ba- sale (in blu). Azan. 40x Utilizzando la colorazione tricromica tutto ciò che è proteico è evidenziato di blu. L'epitelio si deve infatti ancorare ad una matrice molto densa di proteine, essa è chiamata membrana basale (o erroneamente lamina basale). Esistono patologie in cui le proteine che la compongono sono mutate, di conseguenza non sono in grado di fornire sostegno. In questi casi l'epidermide si sfalda, letteralmente, causando l'esposizione del derma ed un alto rischio di infezione. La membrana basale ha un ruolo fondamentale anche nella formazione del carcinoma, cioè del tumore di origine epiteliale. Quando è insito nell'epitelio, la membrana basale funge da barriera che impedisce al tumore di accedere al tessuto connettivale e ai vasi sanguigni, causando metastasizzazione. L'ancoraggio tra le cellule basali e la membrana basale è utile anche per interagire con l'ambiente circostante, infatti alcune proteine funzionano da trasduttori di segnali, sono in grado di innescare cascate di fosforilazioni che inducono trascrizioni di fattori proliferativi (es le integrine funzionano proprio da meccano-trasduttori dei segnali, sono infatti marcatori tumorali perché nel tumore la loro trascrizione viene regolata, e sono induttori delle vie mitotiche). Le giunzioni hanno una distribuzione specifica, non sono disposte lungo i margini cellulari a caso.
Giunzioni Cellulari Epiteliali
2Giunzioni in una cellula epiteliale APICAL Giunzioni occludenti (impermeabilizzazione ) Giunzioni strette OCCLUDING JUNCTION junctional complex CELL-CELL ANCHORING JUNCTIONS Giunzioni di ancoraggio (cellula-cellula) Giunzioni aderenti Collegate ad Actina Desmosomi Collegate a Filamenti intermedi CHANNEL-FORMING JUNCTIONS CELL-MATRIX ANCHORING JUNCTIONS Giunzioni comunicanti (trasferimento di molecole tra cellule adiacenti) Giunzioni Gap BASAL Giunzioni di ancoraggio (Cellula - matrice) Contatti focali Collegate ad Actina Emidesmosomi Collegate a Filamenti intermedi Ipotizziamo che nell'immagine sovrastante sia rappresentato l'epitelio della mucosa intestinale, è monostratificato e le cellule che lo compongono sono di forma rettangolare. Esso è l'esempio migliore per descrivere le giunzioni, in quanto presenta tutte le tipologie che possiamo trovare in un epitelio. La parte rivolta verso la cavità, ovvero lo strato apicale, ha un sistema giunzionale di tipo occludente. Questo vuol dire che vi è un "accollamento" delle membrane citoplasmatiche che sono tenute vicine da proteine, rappresentate in verde. Gli spazi tra una cellula e l'altra sono talmente ristretti da permettere il passaggio esclusivamente a molecole d'acqua e piccoli ioni. Sono necessarie in quanto ricordiamo che l'epitelio intestinale deve assorbire piccole proteine, scinderle in amminoacidi e portarle in circolazione nelle arterie intestinali. Per assorbire tali sostanze gli enteriociti devono utilizzare meccanismi di trasporto attivo, quindi le giunzioni occludenti servono proprio a creare uno strato sigillato verso la cavità. Mano a mano che si va verso lo strato basale delle cellule, sono presenti varie classi di giunzioni ancoranti che servono a tenerle legate tra di loro. Attraverso queste può avvenire il passaggio di piccoli amminoacidi, piccoli peptidi, ioni, sostanze a basso peso molecolare ecc ecc. I complessi proteici delle giunzioni sono tutti connessi con il citoscheletro della cellula, questo collegamento è utile affinché la cellula sappia cosa succede attorno a sé e possa rispondere trasducendo i segnali meccanici ricevuti, in segnali biochimici. Le giunzioni occludenti si trovano a livello apicale, servono per sigillare e sono formate da complessi proteici che creano interazioni molto forti tra di loro in quanto devono resistere a sforzi meccanici.
Proteine delle Giunzioni Occludenti
3citoplasma subapicale dominio apicale della membrana plasmatica proteina intermembranosa canalicoli interstiziali dominio basolaterale della membrana plasmatica Le proteine che le costituiscono sono omodimeriche, cioè la stessa proteina è espressa in entrambe le cellule coinvolte F-actina JAM ine formano adiacenti e. Afadina Nectina Occludina Claudina ZO-1 ZO-2 ZO-3 Via paracellulare Via transcellulare Punto di contatto Le principali proteine che entrano in gioco sono l'occludina e la claudina, le quali formano una sorta di "pinza". Le proteine ZO-1 ZO-2 e ZO-3 costituiscono la basa intracellulare sulla quale l'occludina e la claudina si poggiano (la prof dice che queste ultime sono le uniche proteine da ricordare). La via paracellulare è la zona tra due cellule la quale è completamente chiusa, favorendo la via transcellulare. In queste immagini sono ben visibili le connessioni tra il sistema giunzionale e le fibre del citoscheletro, quali le cheratine, le fibrilline, le actine, i microtubuli ecc. Le giunzioni aderenti si dividono in varie classi che sfruttano diversi tipi di legami.
Giunzioni Aderenti e Molecole di Adesione
4siti che legano Ca2+ fooddo caderine 30000- INTERAZIONI OMOFILICHE domini lg superfamiglia delle Ig (IgSF) recettori per le selectine (CAM simil-mucina) selectine carboidrato INTERAZIONI ETEROFILICHE a B integrine fibronectina Spazio extracellulare Figura 5.19 Molecole di adesione cellulare (CAM). Le proteine di adesione della famiglia delle caderine e della superfamiglia delle immu- noglobuline (IgSF) stabiliscono un legame omotipico (omofilico), in cui interagiscono due molecole identiche appartenenti a due cellule vicine. Il legame che intercorre tra altri tipi di CAM (per esempio selectine e in- tegrine) è considerato eterotipico (eterofilico), poiché le molecole che in- teragiscono non sono identiche. Si noti che le integrine legano proteine della matrice extracellulare (per esempio fibronectina). Per semplicità in questo schema non vengono mostrate le proteine di attacco intracellulari. Tutta questa classe di giunzioni formano quelle che sono chiamate fasce o zonulae aderenti. Osservandole al microscopio elettronico noto un accostamento delle due membrane, ma tutto sommato non è presente una zona scura, quindi gli elettroni possono passare, la cosa cambia osservando i desmosomi. La più importante è quella che sfrutta le caderine, delle proteine utilizzate anche come marcatori tumorali (consideriamo che nei tumori molte giunzioni vengono o represse o over-espresse, perciò la loro de- regolazione provoca un malfunzionamento delle cellule, che perdono anche il loro orientamento). Queste proteine si legano attraverso un legame di tipo ionico, sfruttando il calcio. Sono coinvolte anche le immunoglobuline che funzionano da proteine strutturali legando i propri recettori facendo da ponte tra due cellule. Le giunzioni aderenti presentano anche interazioni eterofiliche per esempio tra recettore e ligiando oppure tra uno zucchero e una proteina. Giunzione occludente Zonula occludens Giunzione intermedia Zonula adhaerens Desmosoma Macula adhaerens - Gap junction (giunzione serrata)
Desmosomi e Emidesmosomi
5I desmosomi sono delle giunzioni aderenti più tenaci proprio perché nel dominio extracellulare esistono delle piattaforme proteiche ad alto peso molecolare. citosol desmogleina (proteine transmembrana) desmoplachina (plaçća intracellulare) interstizio citocheratina (ancorati alla placca) In queste immagini sono visibili le cellule intestinali legate proprio dai desmosomi. Essi servono per dare robustezza al legame tra due cellule vicine, le proteine che partecipano ai desmosomi sono tante, ma dobbiamo ricordare le principali: le desmoplachine formano la piattaforma alla quale si legano le desmogleine, che sono le proteine che sporgono verso l'esterno. Le giunzioni più interessanti sono le giunzioni di ancoraggio alla membrana basale, ovvero gli emidesmosomi. Esse sono costituite dalle integrine, una famiglia di proteine che hanno due funzioni: creare un legame meccanico con le proteine di questa membrana basale e trasdurre dei segnali a livello biochimico. Oltre a fare legami con il citoscheletro, all'interno della cellula legano anche proteine in grado di innescare cascate fosforilatorie, ad esempio le g-protein o sarc-protein.
Ligand CD45 CD4 Antigen LFA-1 Integrin TCR Chemokine receptor Chemokine LAT Lck DAG P P p DAG 6/v P Gga P PLCY1 PLCB P P P PKC IP3 P P S1140 Ca2+ release T758 Zap-70 Talin RAPL S59 Calcineurin RIAM Rap1 GTP Rap1 GDP P Other 4 IP3 ILK CALDAG GEF Trends in Biochemical Sciences 6
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
