Sistema di endomembrane, reticolo endoplasmatico e apparato di Golgi
Documento sul sistema di endomembrane, reticolo endoplasmatico e apparato di Golgi. Il Pdf, di Biologia a livello universitario, descrive la traslocazione e maturazione delle proteine, con schemi e diagrammi esplicativi.
Mostra di più17 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
SISTEMA DI ENDOMEBRANE
La cellula eucariotica possiede numerosi tipi diversi di membrane che delimitano un numero corrispondente di compartimenti. Ogni membrana e compartimento presenta una caratteristica composizione lipidica e proteica che ne determina uno specifico ruolo. E' quindi necessario uno smistamento accurato di lipidi e proteine. Nella cellula quindi esiste un meccanismo di smistamento (sorting) e di direzionamento (targeting) di ogni singola proteina al compartimento di destinazione. L'informazione per il direzionamento risiede generalmente nella proteina stessa sotto forma di sequenze amminoacidiche brevi e distinte. Le sequenza segnale si possono trovare in qualunque punto della proteina: più frequentemente sono localizzate in posizione N-terminale, sottoforma di sequenza transitoria che vengono spesso rimosse nel compartimento di destinazione. Le sequenze segnale e i meccanismi di smistamento sono diversi a seconda della destinazione finale. Tutte le proteine sono sintetizzate sui ribosomi nel citoplasma dove avviene il primo evento di smistamento.
- Una parte di esse rimane nel citoplasma o viene indirizzata ai PALSMIDI, MITOCONDRI, PEROSSISOMI E NUCELO
- Il resto viene indirizzato al RETICOLO ENDOPLASMATICO che fa parte di un vasto sistema definito SISTEMA DI ENDOMEBRANE.
Il SISTEMA DI ENDOMEBRANE comprende: RETICOLO ENDOPLASMATICO, APPARATO DI GOLGI, VACUOLO e MEMBRANA PLASMATICA Tutte le membrane del sistema sono connesse da un traffico vescicolare ANTEROGRADO E RETROGRADO (via di secrezione): sintetizza le proteine che sono destinale alle secrezione, le modifica le assembla le smista e le direzione alle varie destinazioni.Reticolo endoplasmatico Il reticolo endoplasmatico è il compartimento più esteso ed adattabile delle cellule eucariotiche Esso consiste di una rete tridimensionale di tubi continui e sacculi appiattiti che decorrono sotto la membrana plasmatica (RE corticale), attraversano il citoplasma e si connettono all'involucro nucleare (RE tubulare interno). La versatilità di questo compartimento è dovuta alla vasta area superficiale di membrana disponibile per l'incorporazione di proteine (integrali e periferiche) e alla facilità con cui questo sistema può espandersi, ridursi, riorganizzarsi e differenziarsi spazialmente durante il ciclo cellulare e lo sviluppo della cellula. RE tubulare interno è molto mobile rispetto al RE corticale che è in stretta associazione con la membrana plasmatica e con la componente actinica del citoscheletro. Il RE viene suddiviso in:
- RE ruvido ( ribosi adesi alla membrana)
- RE liscio
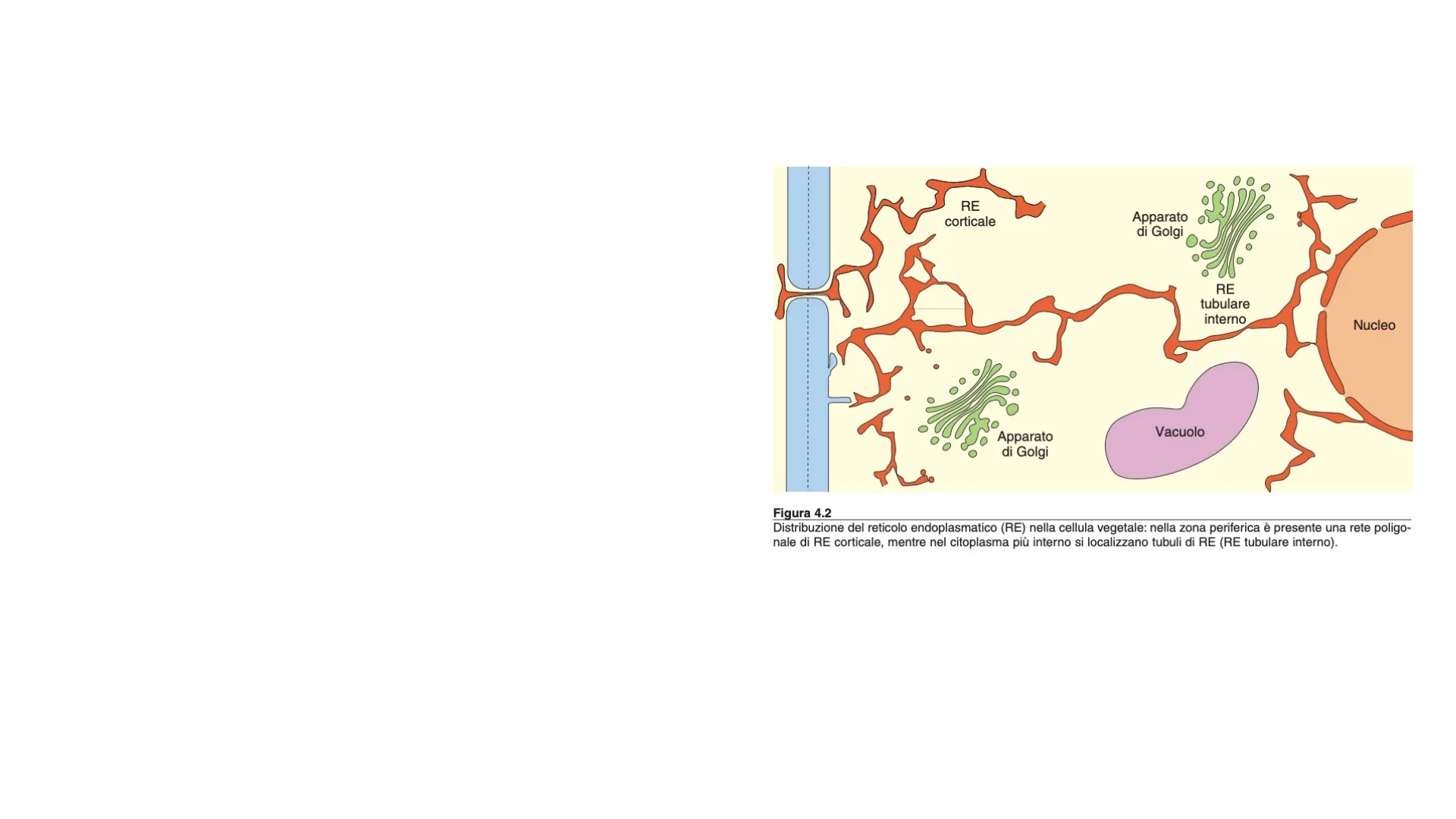
RE corticale Apparato di Golgi RE tubulare interno Nucleo Vacuolo Apparato di Golgi Figura 4.2 Distribuzione del reticolo endoplasmatico (RE) nella cellula vegetale: nella zona periferica è presente una rete poligo- nale di RE corticale, mentre nel citoplasma più interno si localizzano tubuli di RE (RE tubulare interno).Plasmodesma Parete cellularė Vescicola di secrezione Vacuolo .. Apparato di Golgi Corpi proteici RE rugoso RE liscio Actina-F Dominio di aggancio all'actina Oleosine Nucleo Corpi lipidici Il RE è sede della sintesi di proteine e dell'accumulo di alcune proteine di riserva, lipidi di membrana e di lipidi di riserva.
- Proteine di riserva (accumulandosi formano i corpi proteici)
- Trigliceridi (corpi lipidici)
- Oleosine (proteine integrali di membrana): stabilizzano i corpi lipidici e evitano la fusioni tra loro.
Figura 4.4 Alcuni dei domini funzionali attribuiti al reticolo endoplasmatico della cellula vegetale.Nelle piante l'apparato di Golgi è dato dall'insieme di numerose pile, ciascuna costituita da 5-10 cisterne sovrapposte, appiattite al centro e più dilatate ai margini, distribuite in tutto il citoplasma . Ad esse sono associate numerose vescicole. Nella cellula vegetale il numero delle pile varia notevolmente a seconda del tipo di cellula, del suo stadio di sviluppo e della specie a cui essa appartiene. All'interno di ogni pila, le cisterne mostrano una spiccata polarità morfologica basata sulla loro posizione e sulla loro funzione: le cisterne cis sono rivolte verso il RE da cui ricevono, tramite vescicole, i prodotti di sintesi che devono proseguire lungo la via di secrezione. Ad esse seguono le cisterne mediali e le cisterne trans . Associata al lato trans della pila si localizza la regione del "Trans- Golgi network" caratterizzata da strutture tubulari con dilatazioni terminali da cui prendono origine strutture vescicolari. Esso rappresenta il punto di biforcazione a livello del quale avviene lo smistamento delle proteine destinate al vacuolo e di quelle secrete. L'apparato di Golgi è una struttura dinamica che svolge un ruolo chiave nella via di secrezione in quanto nelle sue cisterne si assiste alla maturazione delle proteine neosintetizzate a livello del RE. L'apparato di Golgi è inoltre coinvolto nella sintesi dei glicolipidi di membrana plasmatica e vacuolare e nella sintesi e assemblaggio dei polisaccaridi della matrice parietale. Apparato di Golgi trans-Golgi network cisterna trans cisterna mediale B cisterna cis A C Figura 4.6 A) Visione schematica dell'organizzazione e della posizione reciproca delle cisterne all'interno di una pila dell'appa- rato di Golgi. B) Micrografia al microscopio elettronico delle cisterne trans e del trans-Golgi network in sezione tan- genziale. Sono ben visibili le vescicole in formazione e quelle libere nel citoplasma dopo essersi staccate dalla mem- brana delle cisterne. C) Micrografia al microscopio elettronico di una pila di cisterne disposte come nello schema in A (osservazioni di B. Baldan).TRASLOCAZIONE E MATURAZIONE DELLE PROTEINE NEL RETICOLO ENDOPLASMATICO Ribosoma 3' mRNA 5 Peptide segnale 5 SRP 5' 3 5' 3' 5' 3' Recettore di SRP Peptidasi del segnale Traslocone Proteina Il primo evento di indirizzamento di una proteine alla via di secrezione avviene quando i ribosomi liberi nel citoplasma, a sintesi proteica già iniziata, si associano alla membrana del RE. Tutte le proteine solubili o di membrana destinate alla via di secrezione possiedono all'estremità N-terminale una sequenza aminoacidica, definita peptide segnale, che le indirizza al lume o alla membrana del RE. Le proteine di membrana hanno anche una sequenza supplementare, detta sequenza di arresto del trasferimento, che impedisce la traslocazione completa della catena nascente, mantenendola inserita nella membrana. Due fattori sono importanti per la traslocazione delle proteine nel RE: la particella citosolica che riconosce il peptide segnale (SRP, signal recognition particle) e il recettore di membrana del RE che lega la particella (recettore di SRP). In Figura è possibile seguire le tappe che portano alla traslocazione di una proteina nel lume del RE. SRP si lega alla sequenza del peptide segnale nel polipeptide nascente. La sintesi proteica si arresta temporaneamente e il complesso ribosoma-catena polipeptidica nascente-SRP entra in contatto con il recettore di SRP. Questo riconoscimento media l'aggancio del ribosoma ad un sistema di traslocazione che consente, in concomitanza con la ripresa della sintesi proteica, l'inserimento del polipeptide nascente nel RE. Il peptide segnale viene tagliato da una peptidasi residente nel RE, non appena la catena polipeptidica in allungamento emerge dal sistema di traslocazione. Quest'ultimo, definito traslocone, è costituito da un complesso di polipeptidi che si associano a formare un canale idrofilico di accesso per le proteine.Nel RE la struttura dei polipeptidi neosintetizzati viene modificata in modo che le proteine acquisiscano la corretta conformazione e possano svolgere la propria funzione nel compartimento a cui sono destinate. Gli eventi più frequenti consistono nella N-glicosilazione, formazione di ponti disolfuro, idrossilazione di alcuni residui aminoacidici e assemblaggio di proteine multimeriche. La N-glicosilazione, cioè il legame covalente di un oligosaccaride al gruppo amminico (-NH) della catena laterale di una asparagina, avviene ad opera di una glicosil transferasi che aggancia alla proteina, prima del suo ripiegamento, una catena ramificata costituita da 14 zuccheri. Gli oligosaccaridi vengono modificati nel RE mediante rimozione di alcuni zuccheri, portando alla formazione delle cosiddette catene ad alto mannosio. La funzione principale della N-glicosilazione a garantire un corretto ripiegamento della proteina. Questo processo coinvolge una serie di meccanismi che collettivamente assicurano che solamente le proteine correttamente ripiegate, assemblate e modificate siano fatte proseguire lungo la via di secrezione. Nel lume del RE risiede una serie di enzimi e di chaperonine che accompagnano e assistono la maturazione delle proteine. Una proteina correttamente ripiegata non ha più possibilità di interagire con le chaperonine e viene inviata ai siti di uscita dal RE per essere trasportata all'apparato di Golgi. Invece le proteine difettose, dopo numerosi tentativi di correzione da parte delle chaperonine, vengono trasferite al citoplasma e qui degradate. 1 NH I 1 O Asn X x- O Ser o Thr (A) CO REGIONE CENTRALE (B) OLIGOSACCARIDE COMPLESSO LEGENDA = N-acetilglucosammina (GlcNAc) = mannosio (Man) = galattosio (Gal) = acido N-acetilneuramminico (acido sialico, o NANA) (C) OLIGOSACCARIDE AD ALTO MANNOSIOLe proteine solubili destinate a svolgere la loro funzione nel RE presentano una sequenza molto conservata di quattro aminoacidi K/HDEL (Lys/His-Asp-Glu-Leu), (lisina/istidina, asparagina, glucina, leucina) all'estremità C-terminate. Questa sequenza, detta segnale di ritenzione, e sufficiente a garantire la localizzazione delle proteine residenti nel RE. In realtà e stato dimostrato che alcune proteine, pur possedendo la sequenza di ritenzione, escono dal RE. Esiste infatti un meccanismo di recupero al livello del cis-Golgi dove un recettore di membrana riconosce le proteine con il segnale di ritenzione. Le proteine di RE "fuggite" vengono riciclate all'indietro mediante vescicole del trasporto retrogrado. Quindi, la residenza di una proteina nel RE e il risultato globale di un meccanismo di ritenzione e di recupero selettivo.
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
