Il Muscolo Scheletrico, Parte 2: Fisiologia e Contrazione Muscolare
Documento dall'Università di Ferrara su Il Muscolo Scheletrico, Parte 2. Il Pdf, utile per lo studio universitario di Biologia, esplora la fisiologia del muscolo scheletrico, dettagliando il meccanismo della contrazione muscolare, il ruolo del calcio e i fattori che influenzano la forza muscolare.
Mostra di più15 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Fisiologia del Muscolo Scheletrico
Fisiologia, lezione 10, 17/03/22023
Prof. Viaro
IL MUSCOLO SCHELETRICO, PARTE 2
Il professore riprende da alcuni concetti della lezione precedente.
M-ADP-P.
Fibra
attivata
ADP
Attacco dell'ATP alla miosina,
dissociazione della miosina dall'actina,
idrolisi dell'ATP e recovery stroke
Legame del calcio con la troponina
e modificazione della conformazione
della tropomiosina
ATP
A-M **
Stato rilasciato
energizzato
2
M-ADP-P
‘Ca"
Sito di legame
per la miosina
State
di rigo
6
Stato rilasciato
energizzato
3
Dissociazione
dell'ADP
Formazione del complesso
actina-miosina
(in presenza di calcio)
A-M **- ADP
A-M *- ADP+P
Power
stroke
Dissociazione del fosfato
e generazione di forza
Università di Fefr
Complesso
actina-miosina
senza sviluppo
di forza
4
Contrazione Muscolare e Ciclo dei Colpi di Forza
Contrazione muscolare
Un solo colpo di forza delle teste della miosina
non è abbastanza, ma serve un susseguirsi
ciclico di questi colpi di forza perché ci sia
accorciamento. Importante da ricordare è la
presenza dei sarcomeri (le unità funzionali del
muscolo formate da actina e miosina), i quali,
oltre ad essere in parallelo, sono anche in serie.
Facendo riferimento all'immagine a lato,
vediamo che quando la testa della miosina è
attiva ci sono l'ADP (verde nell'immagine), i
due fosfati P (rossi) e il P separato, quindi
ADP+P. Quando l'ATP è legato come ADP+P
(quindi non è più ATP), vuol dire che l'ATP è
già stato idrolizzato dall'ATPasi che è nel sito
catalitico della testa della miosina.
Legame rotto vuol dire energia, reazione esoergonica.
Quando è sottoforma di ADP+P separati, ma attaccati alla miosina, allora la testa è carica e pronta per scattare.
Questa è una fase di energia.
Successivamente arriva il Calcio (Ca2+), si lega alla troponina e la tropomiosina viene "spazzata via". A questo
punto si legano la miosina e l'actina (come si vede nelle figure in basso dell'immagine). Ora ci sarà il colpo di
forza, detto anche power stroke.
La miosina perde il fosfato P mentre è piegata e rimane solo ADP nella testa. Poi perde anche l'ADP. Quindi
perde sia ADP che P, prima uno poi l'altro, ma sempre durante la "remata". Quando la testa rimane piegata
vuol dire che non c'è alcuna molecola legata e questo si chiama complesso di rigor. Vuol dire che c'è rigidità
muscolare perché c'è già stata la contrazione. La testa della miosina rimane piegata finché non si attacca un
nuovo ATP. Quindi lo stato di rigor c'è sempre (rigor mortis quando a causa della morte non abbiamo l'energia
necessaria per rompere il complesso di rigor). Normalmente lo stato di rigor dura pochissimo e, avendo energia,
un altro ATP si lega e ricomincerà il ciclo dei colpi di forza appena visto.
Alla fine è sempre una questione di ingombro sterico: le molecole si spostano con i vari legami che si formano.
Principio della Leva Molecolare
1PRINCIPIO DELLA LEVA
Uno spostamento di un nanometro (nm) in un punto del fulcro corrisponde a 10nm dall'altra parte della leva
molecolare. Quindi con un piccolo spostamento conformazionale ci si sposta di tanto perché la conformazione
tridimensionale delle molecole è disposta in modo che si sposti in questo modo.
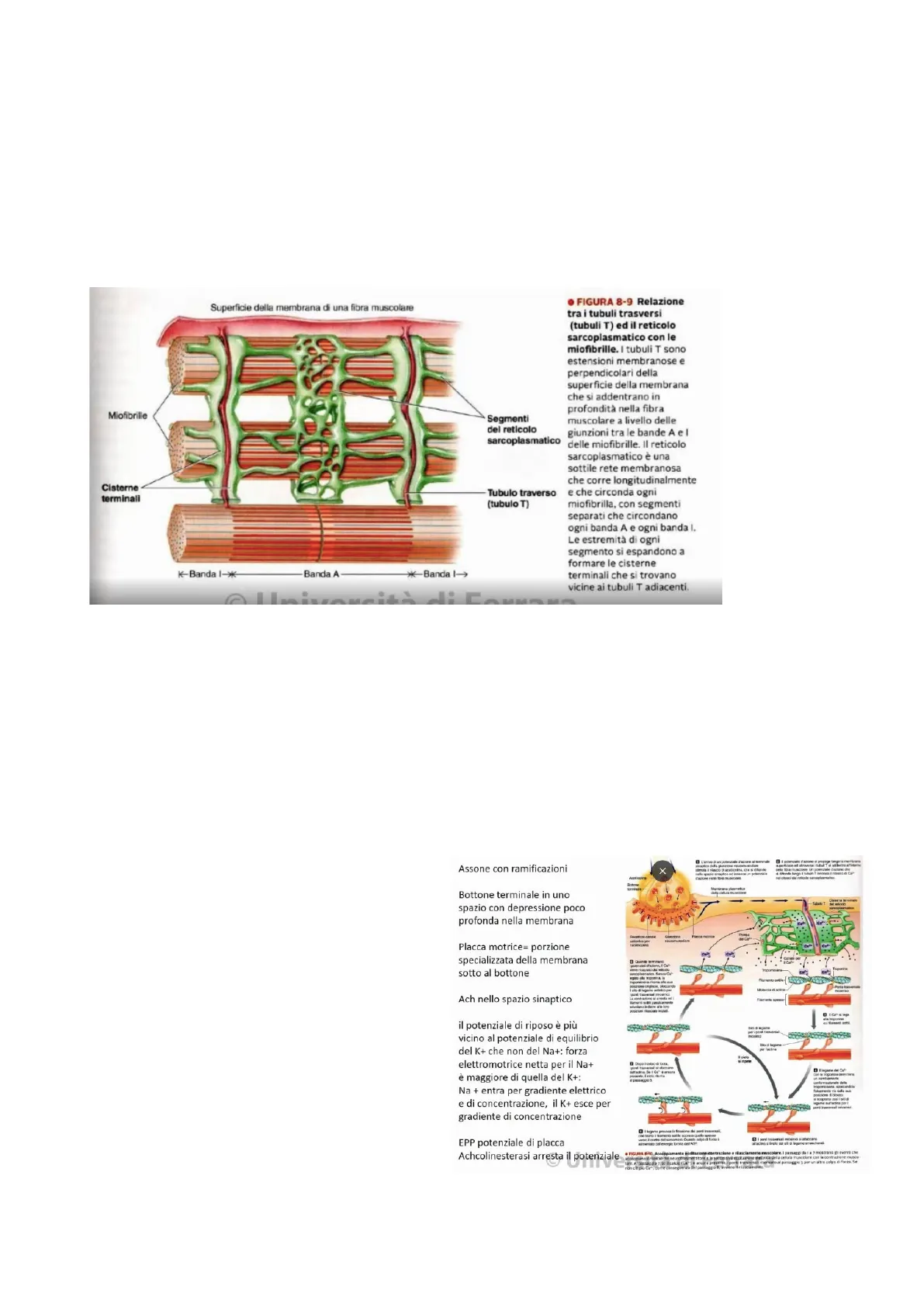
Struttura della Fibra Muscolare
Superficie della membrana di una fibra muscolare
Miofibrille
Segmenti
del reticolo
sarcoplasmatico
Cisterne
terminali
Tubulo traverso
(tubulo T]
K-Banda I -**
Banda A
**- Banda I->
- FIGURA 8-9 Relazione
tra i tubuli trasversi
(tubuli T) ed il reticolo
sarcoplasmatico con le
miofibrille. I tubuli T sono
estensioni membranose e
perpendicolari della
superficie della membrana
che si addentrano in
profondità nella fibra
muscolare a livello delle
giunzioni tra le bande A e I
delle miofibrille. Il reticolo
sarcoplasmatico è una
sottile rete membranosa
che corre longitudinalmente
e che circonda ogni
miofibrilla, con segmenti
separati che circondano
ogni banda A e ogni banda I.
Le estremità di ogni
segmento si espandono a
formare le cisterne
terminali che si trovano
vicine ai tubuli T adiacenti.
Accoppiamento Eccitazione-Contrazione
ACCOPPIAMENTO ECCITAZIONE-CONTRAZIONE
L'accoppiamento eccitazione-contrazione è dovuto al Calcio Ca2+ (il prof dice che questa è una definizione
da sapere). Eccitazione vuol dire che il potenziale d'azione arriva alla giunzione neuromuscolare, (acetilcolina,
giunzione, potenziale postsinaptico di placca che è sempre sopra la soglia ... ). L'eccitazione è una corrente,
invece la contrazione è sempre una cosa meccanica.
Quindi il collegamento tra il segnale elettrico e quello meccanico è dovuto al Ca2+.
Il rilascio del Ca2+ è dovuto alla struttura dei Tubuli T (vedi immagine), ovvero un'invaginazione della
membrana, la quale va giù e risale (come un solco cerebrale). Il potenziale d'azione che corre, grazie a questi
tubuli, va anche dentro le cellule e in profondità (se
Assone con ramificazioni
O Lavivo di un pokonzule d'ozone al se
×
einutico dela gunrice neuromuscolare
nello spazio sinagece at ireesra un powinnae
non ci fossero questi tubuli il potenziale d'azione
Aceticương
Bottone terminale in uno
spazio con depressione poco
profonda nella membrana
rimarrebbe in superficie e non entrerebbe nelle
Placca sofres
cellule muscolari). Quindi il potenziale d'azione
Placca motrice= porzione
specializzata della membrana
...
.
sotto al bottone
(PA) depolarizza anche le parti interne della cellula.
Trocomich
Molecola di acre
Ci sono poi delle cisterne sul reticolo
Ach nello spazio sinaptico
La cort ancre & a testa wo
Flurento apassa
sarcoplasmatico che contengono Ca2+ (la parte
il potenziale di riposo è più
vicino al potenziale di equilibrio
per a pont an
del K+ che non del Na+: forza
elettromotrice netta per il Na+
pordi trapemali u dlaccaro
è maggiore di quella del K+:
presente. i ciclo tierra
Na + entra per gradiente elettrico
e di concentrazione, il K+ esce per
gradiente di concentrazione
verde nell'immagine sopra) e quando sentono il PA
che corre, rilasciano il Ca2+ (viene rilasciato vicino
e ovunque grazie al reticolo sarcoplasmatico).
Concetto importante da ricordare
nella
consultazione delle immagini: solitamente nelle
superficiele ed attraverso i tubul T'ai addiotira affintime
si diffonde lungo il tubulo finnasos i rissois el Cap
Catena berros
Porta Bel
con la tioprina determina
tropomiosona, apostandola
feicaverne va dala qua
posizione di biosoon:
D : line provoca la fessione de ponti kasversal.
the trone a serveres seele sopra a quycho spesso
verso i centrs del soromero, Quesin onipo di forza &
Ø 1 pont bramensal nicunió si attaccano
sone elettrica dela cr
trica della cellula muscolare con la contrazione musco-
EPP potenziale di placca
Achcolinesterasi arresta il potenziale *FIGURA 610 Accoppiamento eccitazione-contrazione e rilasciamente muscolare. I passaggi da 1 a 7 mostrano gi eventi che
Arpaesaggio 70se il calcio (Cat) + ancora presente, i ponti trasversali ritornano al passaggio y per un altro colpo di forza. Se
guenza del passaggio 8: avviene ifriascamento.
Immagine 3
2immagini si vede un solo bottone sinaptico, in realtà un motoneurone che arriva su una cellula muscolare non
arriva solo in un punto, ma può arrivare anche in diversi punti della stessa
cellula (come un braccio con una mano in cui le dita arrivano in punti
Cemponerte
(sarcomeri)
diversi). Quindi la forza dei bottoni più sommarsi. Importante da ricordare è
Cemponer
plastica in serie
mana intracellulare
Carico
nessun connettivo
che sono diverse branche dello stesso motoneurone.
Si accorciano con il colpo di forza
(sarcomeri)
Non si accorciano
(tinina, tendini, ... )
Carico
- FIGURA 8-13 Relazione tra la componente contrattile e
quella elastica in serie nella trasmissione della tensione
muscolare all'osso. La tensione muscolare è trasmessa
all'osso attraverso lo stiramento e la trazione della titina,
tessuto connettivo intracellulare elastico ch
nettre intracellulde elastico che si trova nella
fibra muscolare, e del tendine (che insieme formano la
componente elastica in serie) come risultato
del'accorcamento del sarcomero causato datpatriota cici Ferrara
Immagine 4
Meccanismo della Contrazione e Ruolo delle Proteine
MECCANISMO DELLA CONTRAZIONE
La contrazione avviene finché c'è il Ca2+ che gira
(rilasciato dalle cisterne descritte prima). Quando il
Ca2+ viene tolto, allora finisce la contrazione. Questo
avviene perché il potenziale d'azione non arriva più e
quindi il Ca2+ non viene rilasciato. Il Ca2+ viene
continuamente rilasciato e quello che c'è in circolo
viene poi ricaptato dentro.
Ruolo delle proteine (riferimento immagine 4):
- RYR: rianodina. Quando il potenziale d'azione
(segnale rosso n.4 nell'immagine) arriva nel tubulo
T, attiva il complesso Ryr. - CALSEQUESTRINA: tiene bloccati gli ioni Ca2+ nelle cisterne finché non viene attivato il complesso
Ryr, allora il Ca2+ può uscire.
Il Calcio uscito va quindi alla legarsi alla troponina e la tropomiosina viene "spazzata via". - SERCA: proteina sul reticolo e serve per recuperare il Ca2+.
Rilassamento Muscolare
RILASSAMENTO
Quando non ci sono più impulsi per il movimento, la corteccia non manda più il
potenziale, l'acetilcolina rimane dentro le vescicole e quindi si attica SERCA,
proteina del reticolo che prende il Ca2+ che c'è in giro e lo porta dentro le ;
cisterne dove le calsequestrine lo terranno di nuovo bloccato. Quindi la troponina
non è più legata al Ca2+ e si ripiega, le tre parti globulari si rimettono a triangolo
e la tropomiosina copre i siti della miosina in modo che non
Immagine 5
possa più legarsi.
Tempo di
contrazione
Tempo di
rilasciamento
Scossa
muscolare
Tensione
..
Risposta
contrattile
-
Potenziale di membrana (mV)
30
Potenziale
d'azione
c
-90
1-2 ms
1
50
100
Stimolazione
Tempo (ms)
- FIGURA 8-12 Relazione tra un potenziale d'azione e la
scossa muscolare risultante. La durata del potenziale
d'azione non è disegnata in scala ma è esasperata. Si noti che il
potenziale a riposo di una fibra muscolare scheletrica è -90
milliards /mV\ rispetto a quello di un neurone che è di -70 mV.
Contrazione muscolare
1
PA
Motoneurone
O
Ca
Canali per il Ca2+
Tubulo T
4
ACh
r
Giunzione
neuromuscolare
3
RYR
Cal:
5
Fibra muscolare
SERCA
DHPR
O
Calsequestrina
6
Filamento
di actina
7
Teste
di miosina
Troponina +
tropomiosina
Miofilamenti
3
Nell'immagine 3 si vede lo stesso processo visto in precedenza. C'è del
Ca2+, il colpo di forza ...
Riassumendo, il ciclo si ripete e c'è un susseguirsi di colpi di forza. Mentre
alcune actine e miosine sono legate, altre miosine si riposano e si ricaricano prendendo
ATP che diventa ADP+P e così comincia un nuovo ciclo.
2
Recettori nicotinici
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
