Citologia: cromatina, nucleolo e ciclo cellulare con mitosi
Documento di Appunti sulla citologia, con focus su cromatina, nucleolo e ciclo cellulare. Il Pdf esplora la struttura e la funzione della cromatina, gli stadi di condensazione e l'organizzazione nel nucleo, il nucleolo e il suo ruolo nella biogenesi ribosomiale, il ciclo cellulare, i suoi checkpoint e le fasi della mitosi, per studenti universitari di Biologia.
Mostra di più11 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
La Cromatina
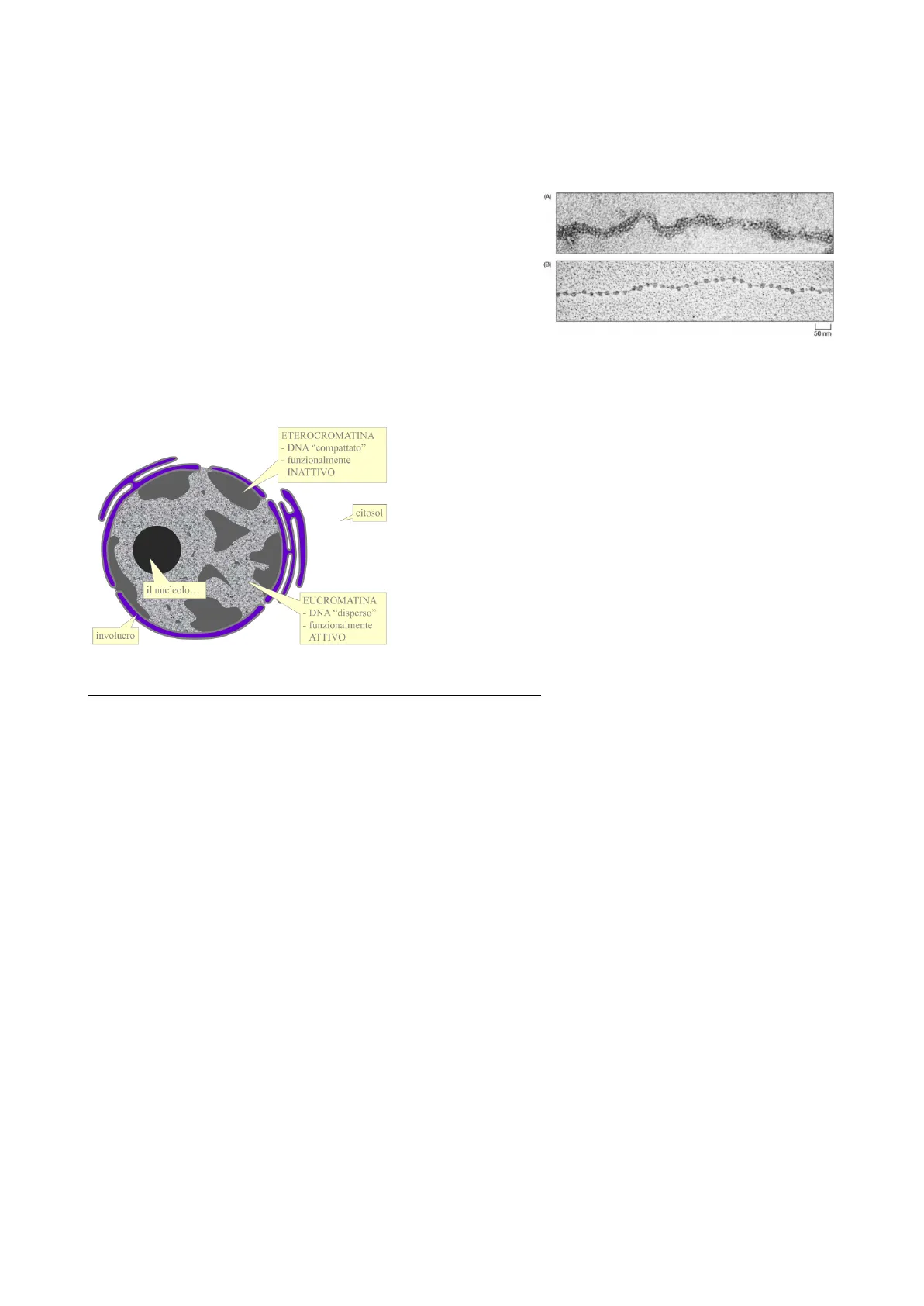
La cromatina è una complessa struttura composta da DNA e (A) proteine che si trova nel nucleo delle cellule eucariotiche. La cromatina si organizza in nucleosomi, unità costituite da un ottamero di proteine istoniche attorno al quale si avvolge il (B) filamento di DNA. Questa struttura assume una forma nota come "collana di perle," che rappresenta l'eucromatina attiva e trascrivibile. Tuttavia, nel nucleo è presente anche un'ampia quantità di cromatina che non è sempre attiva: l'eterocromatina, essendo molto compatta e con un DNA altamente condensato, è generalmente inattiva dal punto di vista trascrizionale. La distribuzione di eucromatina ed eterocromatina all'interno del nucleo riflette l'attività genica della cellula in un determinato momento. Le parti di cromatina che sono utili per la cellula vengono trascritte in base alle necessità specifiche, cambiando continuamente nel tempo, tranne per alcune regioni che rimangono costantemente attive, che specificheremo dopo ...
Nel nucleo è presente anche tanta eterocromatina, come quella nella seguente immagine, che possiede un nucleo con molta eterocromatina elettrondensa poggiata sia sulla lamina subnucleare sia a zolle all'interno del nucleo sparsa tra l'eucromatina.
Stadi della Condensazione della Cromatina
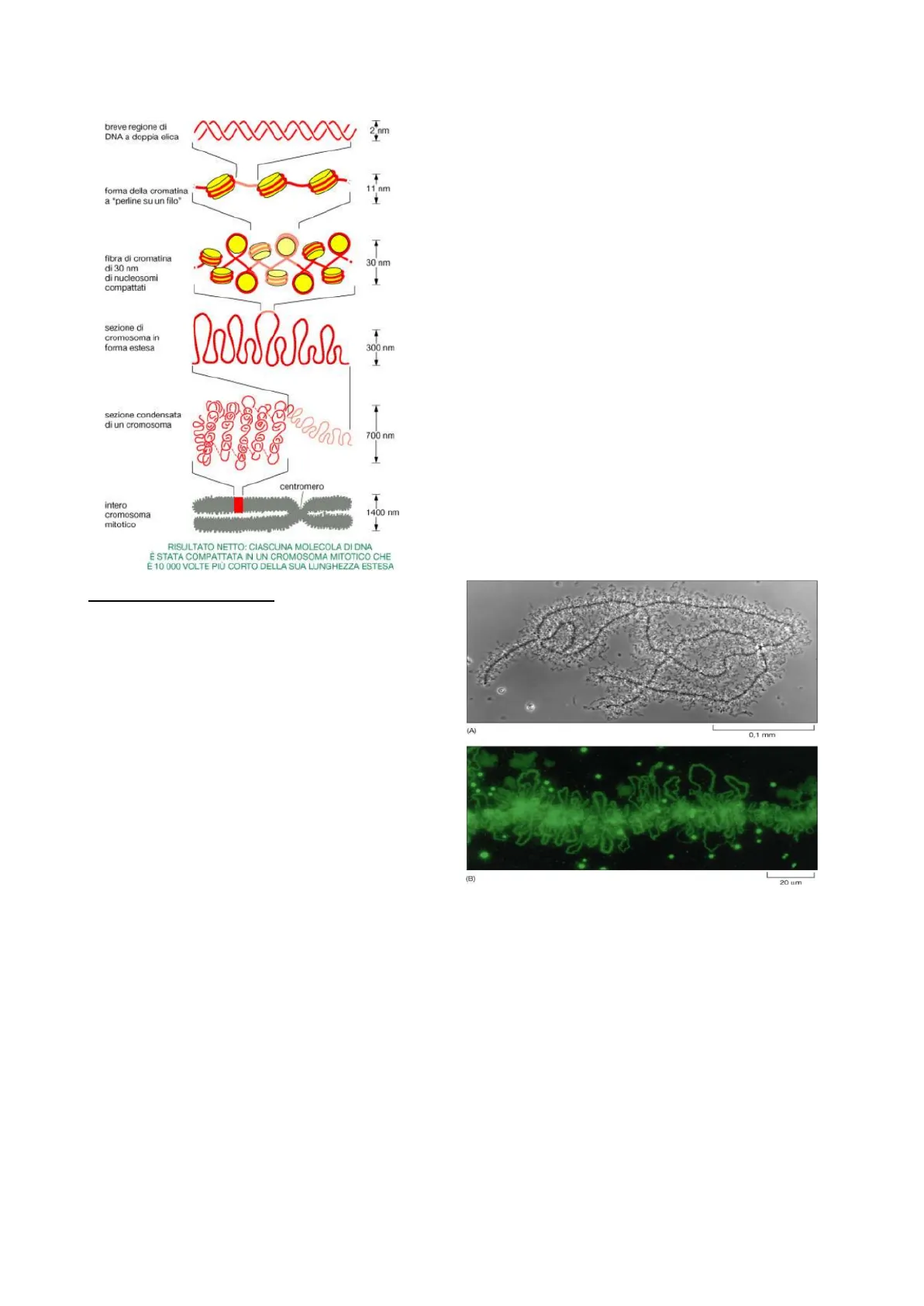
L'eterocromatina si forma per azione dell'istone H1, l'unico istone che non entra a far parte dell'ottamero presente nel nucleosoma; infatti, gli istoni H1 vengono legati dal DNA linker, cioè quel tipo di DNA che congiunge nucleosomi successivi nella collana di perle e abbraccia nucleosomi che si trovano ad una certa distanza nella catena e in questo modo fa sì che il filamento di DNA si ripieghi in una forma che è molto più corta, accorciandosi e aumentando il suo diametro da 11nm a 30nm.
Questo ripiegamento è fondamentale perché segna il passaggio dalla cromatina attiva a cromatina inattiva, cioè eterocromatina non trascrivibile. Tuttavia, anche l'eterocromatina condensata in tal procedimento ha diversi modelli che cercano di spiegare come l'istone H1 interviene e quale tipo di struttura l'eterocromatina da 30nm assume.
Quello che però noi sappiamo è che anche nella versione in cui l'H1 interviene, tutto il DNA non può essere contenuto nel nucleo; dunque, subirà un ulteriore accorciamento e di conseguenza ci aspettiamo che il diametro aumenti ancora di più. Il suo progressivo aumento è positivo poiché rende la struttura meno fragile. La carica negativa del DNA, dovuta ai gruppi fosfato dello scheletro zucchero-fosfato, viene neutralizzata dall'interazione con gli istoni, proteine basiche ricche di lisina e arginina. Questo legame ionico è fondamentale per il primo livello di compattazione del DNA, formando i nucleosomi. Successivamente, ulteriori proteine si associano al complesso DNA-istoni, contribuendo a formare una struttura più condensata, la fibra di 30nm. Infine, un'impalcatura proteica, lo scaffold, permette alla cromatina di organizzarsi in domini topologicamente associati e di formare i cromosomi, strutture altamente condensate essenziali per la segregazione dei cromatidi durante la mitosi. Questo complesso processo di compattazione del DNA è strettamente regolato e garantisce una corretta trasmissione del materiale genetico alle cellule figlie.
Al momento non abbiamo informazioni sufficienti per descrivere dettagliatamente la condensazione del DNA, ma conosciamo la sua forma più condensata, cioè l'eterocromatina. Infatti, se la trattiamo con DNA polimerasi ed RNA polimerasi o estrazione con detergente in modo tale da togliere proteine scaffold osserviamo di passaggio in passaggio ciò che rimane in questi processi.
La cromatina da 300nm, ottenuta mediante il trattamento dell'eterocromatina e l'estrazione delle proteine scaffold, presenta una caratteristica organizzazione ad anse radiali. Queste anse corrispondono a segmenti di DNA che si estendono radialmente da un nucleo proteico centrale, il quale fornisce il supporto strutturale necessario per mantenere questa conformazione spaziale. La struttura ad anse radiali è fondamentale per l'ulteriore compattazione della cromatina e per l'organizzazione tridimensionale del genoma all'interno del nucleo.
Regioni AT Ripetute e Scaffold Proteico
Regioni AT ripetute: La presenza di uno scaffold proteico (oltre a quello costituito dagli istoni) permette al filamento di spiralizzarsi in una struttura da 300nm e successivamente da 700nm. Le anse di cromatina partono e tornano ad agganciarsi in punti contigui di questa impalcatura proteica, tramite brevi tratti di DNA ricco di AT, noti come regioni di attacco allo scaffold. Quindi il filamento di DNA del cromosoma è un filamento continuo, con piccole porzioni aderenti all'impalcatura proteica e grandi porzioni espanse in anse laterali che costituiscono precisi territori cromatinici.
Domande e Risposte sulla Cromatina
Domanda: Le proteine scaffold si legano soltanto alle AT ripetute o anche a qualcos'altro?
Risposta: No, gli istoni sono già impegnati nel loro lavoro, a cambiare il modo in cui il filamento di DNA si associa per fare i nucleosomi, poi interviene l'ultimo istone H1 che ripiega il tutto, passiamo dalla forma da 11nm di eucromatina a eterocromatina. Per far sì che il DNA sia contenuto all'interno del nucleo, è necessario che la cromatina subisca vari ripiegamenti: il primo ripiegamento avviene grazie all'H1, che prende la collana di perle e forma l'eterocromatina da 30nm. Ogni volta che su questo filamento ci sono delle sequenze AT ripetute queste si vanno ad associare a delle proteine non istoniche e non necessariamente basiche, che li legano a questi domini attivi, le anse radiali che si allontanano e poi si riavvicinano allo scaffold proteico.
Domanda: Queste proteine hanno un nome?
Risposta: No, perché è una famiglia molto ampia come quella degli istoni; dunque, le identifichiamo semplicemente nelle proteine dello scaffold.
La cromatina con diametro di 300nm non è l'ultimo stadio di ripiegamento, in quanto è possibile passare ad una struttura più condensata avente un diametro di 700nm, che rappresenta il diametro dei singoli cromatidi. Dunque, in questa struttura la cromatina condensata forma i cromatidi che poi si uniscono per formare il cromosoma. Questa forma da 700nm non è sempre presente nella cellula, ma lo è quando deve affrontare una divisione cellulare, cioè durante la duplicazione del DNA.
Dunque, nel nucleo di una cellula qualsiasi, durante l'interfase, noi troviamo una parte di eterocromatina da 30nm, da 300nm e una parte di eucromatina da 11nm. Se noi vogliamo sapere quanto misura la fibra della cromatina che stiamo osservando dobbiamo utilizzare il microscopio elettronico.
Durante la metafase, i cromosomi da 1400nm sono visibili come strutture discrete e altamente condensate, formate da due cromatidi fratelli (da 700nm ciascuno) uniti al centromero. I cromosomi avendo una dimensione che va oltre il potere di risoluzione del microscopio ottico, è possibile analizzarli quando la cellula va in mitosi e tutta la cromatina si condensa in cromosomi, che sono meno fragili e più facilmente divisibili tra le due cellule figlie.
Il Cariotipo
Il cariotipo viene analizzato attraverso un esame microscopico delle cellule in fase di divisione. La parola "cariotipo" deriva dal greco "karyon," che significa "nucleo," e "typos," che significa "tipo" o "modello." Quindi, etimologicamente, "cariotipo" si riferisce al "tipo di nucleo."
Se noi stiamo facendo il cariotipo significa che quella cellula si sta dividendo e che tutta la cromatina della cellula è diventata eterocromatina e non c'è più il nucleo morfologicamente visibile con il suo involucro nucleare, membrana esterna, interna, lamina nucleare, complesso del poro, ecc ... perché durante la mitosi l'involucro e la lamina si dissolvono e i cromosomi li troviamo sparsi nel citoplasma.
Esempio: facendo il prelievo del sangue con conseguente estrazione, ciò che osserveremo sul vetrino ci darà la possibilità di vedere il numero di cromosomi; infatti, è anche possibile riconoscere delle malattie cromosomiche, come la trisomia 21, ovvero la sindrome di Down, e posso vederne anche la forma e la dimensione ed eventualmente visualizzare qualche problematica avvenuta nella duplicazione del DNA.
L'Analisi del Cariotipo
Avviene attraverso il prelievo di sangue e prevede vari passaggi: Innanzitutto, si effettuano prelievi di sangue dai quali si isolano i leucociti, poiché gli eritrociti non possiedono un nucleo e non consentirebbero una visualizzazione adeguata dei cromosomi. Le cellule leucocitaria vengono poi coltivate in un ambiente favorevole, dove si somministrano fattori di crescita o altre molecole in grado di stimolare la mitosi. Dopo un periodo di incubazione di circa un giorno, si utilizza il "colcemid", un farmaco antitumorale che agisce bloccando le cellule in metafase. Questo avviene perché il colcemid inibisce la formazione delle fibre del fuso mitotico, impedendo la separazione dei cromosomi. Una volta bloccate in metafase, le cellule vengono estratte e preparate per l'analisi. Grazie a un software specializzato, è possibile visualizzare i cromosomi, che appaiono colorati e distribuiti nel citoplasma, facilitando così l'osservazione e la successiva analisi del cariotipo.
Tipi di Cromatina
Sappiamo che nel nucleo l'eucromatina è diversa da cellula a cellula, ma anche all'interno della cellula si distingue nei suoi diversi momenti di vita.
Nella cellula è presente una parte di eucromatina, comune a tutte le cellule, che è sempre trascrivibile, che va a costituire i geni housekeeping; un'altra parte di eucromatina risulta diversa di cellula in cellula; infatti, in alcune si presenta come eu- e in altre come etero -.
Esempio: Gli epatociti, le cellule del fegato, sono specializzati nella detossificazione e, per svolgere questa funzione, necessitano di esprimere una serie di enzimi del citocromo P450. Al contrario, altre cellule, come quelle dei polmoni, del sangue o delle fibre muscolari, non hanno bisogno di questi enzimi per le loro funzioni specifiche. Di conseguenza, i geni che codificano per gli enzimi del citocromo P450 sono attivi negli epatociti, mentre in altre cellule rimangono silenti e fanno parte dell'eterocromatina. Questo fenomeno dimostra come la regolazione genica consente a cellule diverse di esprimere specifici profili di geni, ottimizzando così le loro funzioni specializzate.
Esempio di Cellule con Eucromatina Diversa
Domanda d'esame: fate qualche esempio di cellule che possono avere eucromatina con geni trascritti diversi.
Risposta: L'esempio più semplice ma non del tutto corretto sono gli eritrociti, che non hanno il nucleo, perché il loro scopo principale è produrre tante globine per produrre tanta emoglobina per portare l'ossigeno attraverso il torrente circolatorio negli stretti corporei.
Sebbene siano privi di nucleo, il loro citoplasma è ricco di mRNA per la produzione di globina, l'unità strutturale dell'emoglobina. Questo mRNA, derivante dalla trascrizione dei geni per le globine, consente agli eritrociti di sintetizzare abbondantemente emoglobina, il che è essenziale per il trasporto dell'ossigeno nel sangue. In questo caso, la presenza di mRNA consente comunque alla cellula di svolgere la sua funzione primaria, nonostante l'assenza del nucleo. In queste cellule il gene della globina sarà eterocromatina, ciò significa che esiste una parte di eterocromatina che può essere decondensata e può diventare eucromatina in base alle condizioni fisiologiche della cellula.
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
