Bioenergetics: Free Energy, ATP, and Oxidation of Carbon Fuels
Document from University about Bioenergetics: Free Energy, ATP, and Oxidation of Carbon Fuels. The Pdf explores key concepts like enthalpy, entropy, and free energy in biology, detailing carbon fuel oxidation and ATP's role. It is suitable for University students studying Biology.
See more22 Pages

Unlock the full PDF for free
Sign up to get full access to the document and start transforming it with AI.
Preview
INTRODUCTION
Bioenergetics -> quantitative study of 1) energy transductions (= conversion of one form of energy into another) taking place in living cells and of 2) chemical processes featuring these transductions
- quantitative observations concerning the changes of different energy forms led to the development of thermodynamics, as well as to the formulation of its laws -> thermodynamic quantities describing energy changes during chemical reactions were defined.
- Enthalpy (H) = heat content of a reacting system, keeping constant pressure -> during a chemical reaction, heat is released (exothermic process) when the heat content of the products is less than that of the reactants, being enthalpy change a negative value (AH<0) # reacting systems absorbing heat from the surroundings are defined as endothermic and are associated to AH>0 Entropy (S) = measure of system disorder/randomness -> reaction yielding products that are less complex and more disordered compared with the reactants proceed with an entropy gain (AS>0).
- Free energy (G) (from J. Willard Gibbs) = amount of energy capable of doing work, associated with a reaction conducted at constant T and P -> reactions proceeding with free energy release are defined exergonic, thus displaying a negative (< 0) AG value # in endergonic reactions, the reacting system gains free energy and AG > 0.
FREE ENERGY
Free energy change/variation (AG) is related with reactions/processes occurring within closed systems (those one exchanging heat, but not matter, with surroundings) at fixed temperature and pressure -> these conditions co-exist within biological systems AG value associated with chemical reactions occurring within cells allows the prediction of 1) their direction + 2) the equilibrium position + 3) the amount of the theoretical performed work.
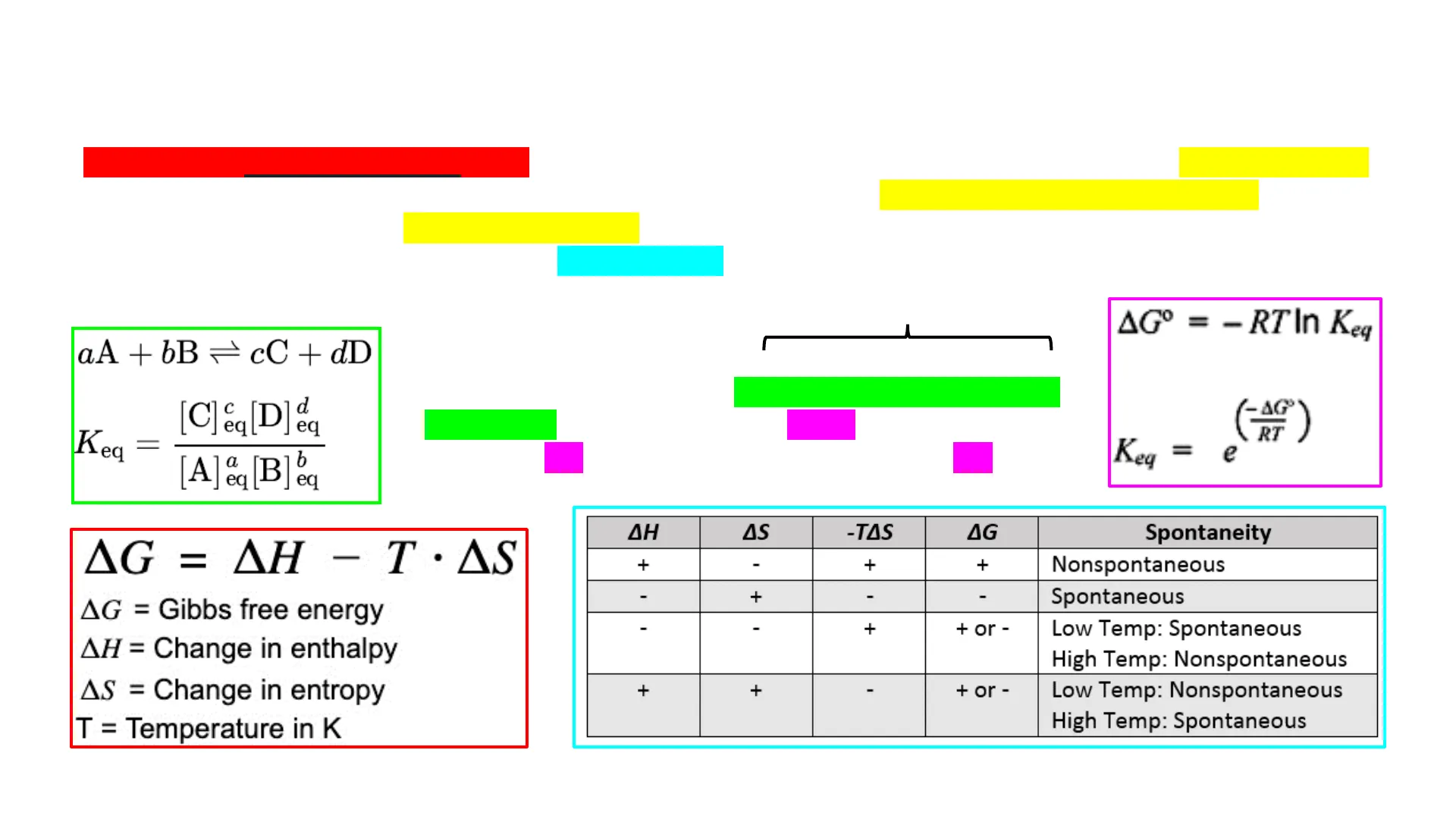
... + 1M starting concentrations of all components aA + bB=cC + dD [C] &q[D]e Keq = [A] & [B]eq Under standard conditions `(P = 1atm + T = 273K and marked with """) and upon reaching the point of equilibrium, it is possible to relate the equilibrium constant Ked with the free energy change eg (AG) AGº = - RTIn K AG Keq = e AG = AH - T . AS AG = Gibbs free energy AH = Change in enthalpy AS = Change in entropy T = Temperature in K
| ΔΗ | AS | -TAS | ΔG | Spontaneity |
|---|---|---|---|---|
| + | - | + | + | Nonspontaneous |
| - | + | - | - | Spontaneous |
| - | - | + | + or - | Low Temp: Spontaneous High Temp: Nonspontaneous |
| + | + | - | + or - | Low Temp: Nonspontaneous High Temp: Spontaneous |
FREE ENERGY RELATIONSHIPS
The standard free-energy change (AGO) of a chemical reaction simply represents an alternative mathematical way of expressing its equilibrium constant (Keg) there is a relationship between this 2 quantities
| AG'® | K'e | (KJ/mol) | (kcal/mol)ª |
|---|---|---|---|
| 103 | -17.1 | -4.1 | |
| 102 | -11.4 | -2.7 | |
| 101 | -5.7 | -1.4 | |
| 1 | 0.0 | 0.0 | |
| 10-1 | 5.7 | 1.4 | |
| 10-2 | 11.4 | 2.7 | |
| 10-3 | 17.1 | 4.1 | |
| 10-4 | 22.8 | 5.5 | |
| 10-5 | 28.5 | 6.8 | |
| 10-6 | 34.2 | 8.2 |
Since there is an exponential relationship between AGO and Keg, relatively small changes in AGº correspond to large changes in Keg AGº = - RTIn Ke G Keq = @
| When Keq is | AG'· is | Starting with all components at 1 M, the reaction ... |
|---|---|---|
| >1.0 | negative | proceeds forward |
| 1.0 | zero | is at equilibrium |
| <1.0 | positive | proceeds in reverse |
FREE ENERGY AND REACTION SPONTANEITY
AG (or AGº) = the difference between G content of the products and G content of the reactants -> AG < 0 is referred to products containing less free energy than the reactants and the reaction will proceed spontaneously # AG < 0 means that the reaction products contain more free energy than the reactants. EXERGONIC REACTION: AG < O ENDERGONIC REACTION: AG > O Reaction is spontaneous Reaction is not spontaneous
Gibbs Free Energy Release
Energy is released reactants AG < 0 products Time
Gibbs Free Energy Added
Energy is added products AG> 0 reactants Time
BIOCHEMICAL REACTIONS
Within cell, thousands of relevant (when an available substrate is converted to a useful product) + specific + enzyme-catalyzed reactions are carried out -> transformation of nutrients into amino acids, nucleotides or lipids + energy extraction from fuels (glucose or fatty acids) by oxidation + polymerization of monomeric subunits (monosaccharides, amino acids or nucleotides) into macromolecules (polysaccharides, proteins and nucleic acids).
- internal rearrangements, isomerizations, and eliminations
- group transfer reactions
- reactions that make or break C-C bonds
- free-radical reactions
- oxidation- reductions
BIOCHEMICAL REACTION MECHANISMS
Covalent Bond Cleavage
Covalent bonds (= shared electron pairs) can be broken by generating a homolytic cleavage, (unpaired electron = radical formation + less common) or a heterolytic cleavage (retention of both bonding electrons = ions formation + more common).
- Homolytic cleavage -C-H =- C + H - Carbon radical 1 -C-C- -C + ℃- 1 - Carbon radicals
- Heterolytic cleavage -C-H I 1 -C:" 1 + H+ Carbanion Proton -C-H I 1 -C+ 1 + H :- Carbocation Hydride - - -C-C- 1L -C:" 1 + + C- Carbanion Carbocation
Nucleophile and Electrophile Interactions
Biochemical reactions include interactions between nucleophile and electrophile species → functional groups rich in electrons and capable of donating them + electron-deficient functional groups that seek negative charged or polarized centers.
| Nucleophiles | Electrophiles |
|---|---|
| :R 0: Negatively charged oxygen (as in an unprotonated hydroxyl group or an ionized carboxylic acid) | R + -CT Carbanion |
| ·S: Negatively charged sulfhydryl | -N Uncharged amine group |
| R TO-P= =0 HN Phosphorus of a phosphate group | Imidazole |
| -: R H-0: H+ Hydroxide ion | Proton |
| Carbon atom of a carbonyl group (the more electronegative oxygen of the carbonyl group pulls electrons away from the carbon) | H Protonated imine group (activated for nucleophilic attack at the carbon by protonation of the imine) |
| N: -C. Hatom 1 I I |
BIOCHEMICAL GROUP TRANSFER REACTIONS
group transfer reactions
O 5+ P-O- δ- O Enzymes catalyzing phosphoryl group transfers, using ATP as donor, are called kinases (i.e. hexokinase, moving a phosphoryl group from ATP to glucose)
O O O= Adenosine HO-P-O-P-O-P-O- HÖ-R - I Glucose 0 O ATP O Adenosine -0-P-O-P-O" + "O-P-O-R 1 1 ADP I -O 0 Glucose 6-phosphate, a phosphate ester Metabolic pathways are often characterized by phosphoryl group (-PO22-) transfers -> it represents a good leaving group that, once attached to a metabolic substrate, is able to activate the intermediate for subsequent reaction -> among the better leaving groups, it participate to hundreds of metabolic reactions. Oxygen (O) is more electronegative than phosphorus (P) -> unbalanced electron sharing = P bears a partial positive charge (o+), acting as an electrophile species -> in many metabolic reactions, a phosphoryl group is transferred from ATP to an alcohol or to a carboxylic acid, thus forming a phosphate ester or a mixed anhydride, respectively.
BIOCHEMICAL REDOX REACTIONS
redox reactions In biochemical molecules, carbon atoms can display five different oxidation states and transitions between these states occur during oxidation-reduction (redox) reactions featuring metabolism -> biological oxidations = e- and H+ loss = dehydrogenation reactions catalyzed by dehydrogenase enzyme.
- CH2 -CH3 Alkane
- CH2-CH2OH Alcohol
- O -CH2-C Aldehyde (ketone) H(R)
- O -CH2-C Carboxylic acid OH
- O=C=0 Carbon dioxide
the oxidation of an electron donor is always associated with the reduction of an electron acceptor -> oxidation reactions generally release energy and living cells obtain the needed energy by oxidizing metabolic fuels such as carbohydrates or fat -> during these (catabolic) energy-yielding processes, electrons are transferred from fuel molecules to oxygen (O2), by using a series of electron carriers. Oxygen display a great electron affinity -> electron- transfer process is highly exergonic, thus providing the energy for ATP synthesis = the central goal of catabolism
OH 2H+ + 2e O O I - CH3-CH-C CH3-C-C 1 1 2H+ + 2e- Lactate lactate dehydrogenase Pyruvate
THE ROLE OF ATP
Within biological reactions (metabolism), ATP plays a fundamental role as the energy currency that links catabolismo and anabolism -> cells of heterotrophic organisms obtain "chemical" energy through catabolic reactions (usually exergonic) of nutrient (complex) molecules.
-O-P-O-P-O-P-O- Adenosine - H-0: O 0- O ATP4- H 0 O 0 1 HO-P-O-P-O- Adenosine -O-P-OH hydrolysis, with relief of charge repulsion O 0- ADP2- 0- P 2 resonance stabilization 8- 3- 0 8-0- P - 0 8- H+ O / ATP4- + H20 - ADP3- + HPO2- + H+ AG'º = - 30.5 KJ/mol This energy is used to synthetize ATP (ADP + Pi) that, in turn, undergoes hydrolysis (accompanied with a wide free energy release) to provide chemical energy to allow cell to carry on endergonic processes featuring 1) anabolism (= synthesis of metabolic O H+ + "O-P-O-P-O- I 1 0- 0 Adenosine ADP3- ionization = intermediates and macromolecules starting from smaller precursors) + 2) transport of substances across membranes against concentration gradients + 3) mechanical motion.
THE ROLE OF ATP IN METABOLISM
Within biological reactions (metabolism), ATP plays a fundamental role as the energy currency that links catabolismoand anabolism -> cells of heterotrophic organisms obtain "chemical" energy through catabolic reactions (usually exergonic) of nutrient (complex) molecules.
O C O -O-P-O-P-O-P-O- Adenosine - H-0: O O ATP4- ー エ O O O 1 HO-P-O-P-O- Adenosine -O-P-OH hydrolysis, with relief of charge repulsion O 0- ADP2- 0- ionization 2 resonance stabilization O O 8- 3- 0 H+ + "O-P-O-P-O- Adenosine 1 0 ADP3- ATP4- + H20 -> ADP3- + HPO2 + H+ AG'º = - 30.5 KJ/mol Chemical basis for the relatively large and negative standard free energy of ATP hydrolysis = cleavage of the terminal (one out of three) negatively charged phosphoryl grou p bond to the rest of the molecule relieves some of the internal electrostatic repulsion + released P can be stabilized by the formation of several resonance forms 8-0- P - 0 8- H+ O=2-0 P -C
THE ROLE OF ATP + OTHER PHOSPHORYLATED COMPOUNDS
| AG'- | Compounds and Acetyl-CoA | (KJ/mol) | (kcal/mol) |
|---|---|---|---|
| Phosphoenolpyruvate | -61.9 | -14.8 | |
| 1,3-Bisphosphoglycerate (-> 3-phosphoglycerate + Pi) | -49.3 | -11.8 | |
| Phosphocreatine | -43.0 | -10.3 | |
| ADP ( -> AMP + P;) | -32.8 | -7.8 | |
| ATP ( -> ADP + P;) | -30.5 | -7.3 | |
| ATP (-> AMP + PPi) | -45.6 | -10.9 | |
| AMP ( -> adenosine + Pi) | -14.2 | -3.4 | |
| PP:(-> 2Pi) | -19.2 | -4.0 | |
| Glucose 3-phosphate | -20.9 | -5.0 | |
| Fructose 6-phosphate | -15.9 | -3.8 | |
| Glucose 6-phosphate | -13.8 | -3.3 | |
| Glycerol 3-phosphate | -9.2 | -2.2 | |
| Acetyl-CoA | -31.4 | -7.5 |
In the cytosol, Mg2+ ions usually binds to both ATP and ADP molecules, in order to stabilize negative charges of phophoryl groups -> in most enzymatically catalyzed reactions involving ATP as phosphoryl group donor molecule, the substrate is not ATP but MgATP2 -.
O O O= TO-P-O-P-O-P-O- Adenosine O O MgATP2- Mg 2+ O C -O-P-O-P-O- Adenosine O O- MgADP- g2+ Mg- In addition to ATP, significantly large, negative free energy values are reported for the hydrolysis of a wide variety of biologically important phosphorylated compounds -> this is due to the several resonance forms available to phosphoryl group (Pi), stabilizing this product that is generated following the associated phosphate-releasing reactions -> i.e. glycolysis intermediates + other important substrates.
Can’t find what you’re looking for?
Explore more topics in the Algor library or create your own materials with AI.
