Tema 1: Ácidos Nucleicos, Apuntes de Biología para Universidad
Documento de Universidad sobre Tema 1: Ácidos Nucleicos. El Pdf explora los ácidos nucleicos, bases nitrogenadas, nucleósidos y nucleótidos, así como la inhibición de la transcripción y maduración del RNA. Es un material de Biología para estudiantes universitarios.
Ver más58 páginas


Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Nucleótidos y Ácidos Nucleicos
Todos los tipos de células contienen un gran número de nucleótidos que son extremadamente importantes en diversas funciones.
La función más significativa de estos nucleótidos es que constituyen la unidad monomérica de los ácidos nucleicos (DNA y RNA). Sin embargo también podemos encontrar nucleótidos que actúan de reserva energética (ATP), como segundos mensajeros en los mecanismos de acción de hormonas y neurotransmisores (AMPc), o el ADP que actúa interviniendo en el mecanismo de agregación plaquetaria y coagulación sanguínea, así como en otra serie de actividades dentro de las vías metabólicas.
La concentración de estos nucleótidos en la célula varia desde concentraciones mM para los ribonucleótidos (RNA) hasta uM para los desoxirribonucleicodos (DNA), siendo el más abundante el ATP. Si bien, la mayoría de las células podrían sintetizar de "novo" los nucleotidos necesarios, lo cierto es que al ser un proceso con un alto gasto de energía, con frecuencia se siguen vías de recuperación de purinas y pirimidinas libres.
Bases Nitrogenadas
Cuando los ácidos nucleicos sufren una hidrólisis total se produce una liberación de sus distintas unidades estructurales; las bases puricas y pirimidímicas, la D-ribosa o la 2-desoxi-D-ribosa (RNA o DNA) y el ácido fosfórico. Las combinaciones de estos componentes son los nucleósidos (base + pentosa) y nucleótidos (base + pentosa + 1, 2 ó 3 grupos fosfato).
Purinas
Las bases púricas derivan de la purina que es un compuesto formado por un anillo pirimidina y un anillo imidazol. Las purinas más importantes son la adenina (A) y la guanina (G). Otras bases púricas naturales son la hipoxantina, la xantina y el ácido úrico y también podemos encontrar una serie de bases modificadas (generalmente con grupos metilo e hidroximetilo) en los RNA transferentes (tRNA).
NHO O - N N HN" CH CH HC C 0 - N H2N N H Adenina Guanina PURINAS
Pirimidinas
Las bases pirimidímicas son derivados de la pirimidina. Son la citosina (presente en el DNA y RNA), la timina (en el DNA) y el uracilo (en el RNA), además de una pequeña proporción de formas metiladas o hidroximetiladas que forman parte de los tRNA.
NH2 O=C CH3 Z CH HN HN CH 0 CH CH CH 0 N 0 N N H H H Citosina Timina (ADN) Uracilo (ARN) PIRIMIDINAS N H2
Nucleósidos
Un nucleósido es una estructura en la que el carbono uno (C1) de la pentosa reacciona mediante enlace glucosídico con el nitrógeno nueve (N9) de la base si es una purina o con el nitrógeno uno (N1) si la base es pirimidina.
Los ribonucleósidos principales son la Adenosina, Guanosina, Uridina y Citidina, todos ellos derivados de la D- ribosa. Los desoxinucleósidos más importantes son la Desoxiadenosina, Desoxiguanosina, Desoxitimidina y Desoxicitidina que derivan de la 2-desoxi-D-ribosa.
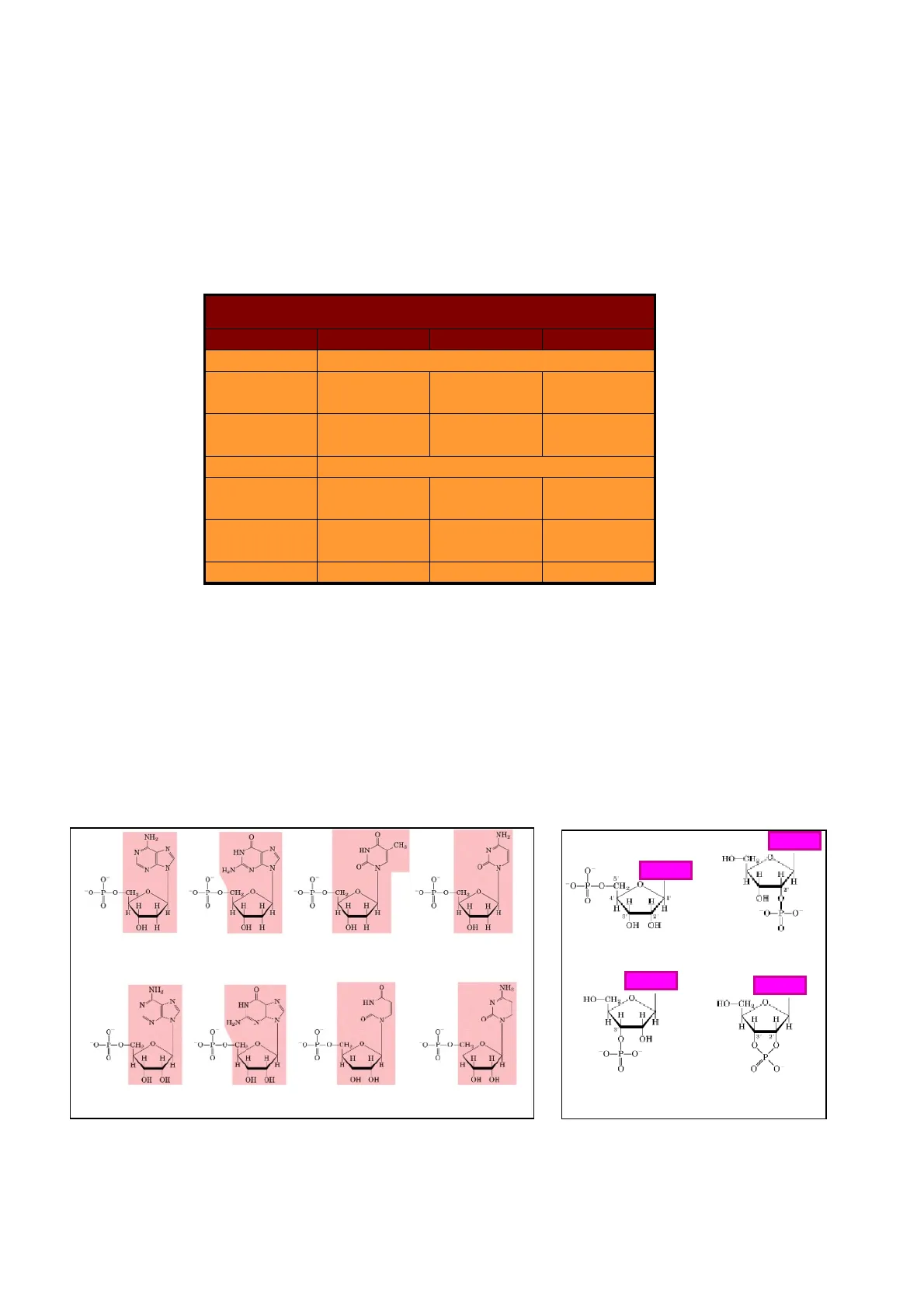
Nomenclatura de Nucleótidos y Ácidos Nucleicos
| Base | Nucleósido | Nucleótido | Ácido nucleico |
| Purinas | |||
| Adenina | Adenosina | Adenilato | RNA |
| Desoxiadenosina | Dexosiadenilato | DNA | |
| Guanina | Guanosina | Guanilato | RNA |
| Desoxiguanosina | Desoxiguanilato | DNA | |
| Pirimidinas | |||
| Citosina | Citidina | Citidilato | RNA |
| Desoxicitidina | DNA | ||
| Timina | Timidina o desoxitimidina | Timidilato o desoxitimidilato | DNA |
| Uracilo | Uridina | Uridilato | RNA |
Nucleótidos
La esterificación de los nucleósidos con ácido fosfórico da lugar a los nucleótidos. El punto de unión más frecuente para el grupo fosfato es el carbono cinco (C5) de la pentosa. Si el azúcar es D-ribosa lo que se obtienen son 5'-ribonucleótidos (AMP, GMP, UMP y CMP) o si es 2-desoxi-ribosa se obtienen 5'-desoxirribonucleicos (dAMP, dGMP, dTMP y dCMP).
NH2 C 0 NH2 CH HN N N HN Z H.N 2 0 0 O O-P O 0 -CH2 O-P-O-CH- -CH 0 H H H H H H H H H H H H 2 OH H OH H OH H OH H Desoxiadenosina Desoxiguanosina Desoxitimidina Desoxicitidina Adenosina 5'-monofosfato Adenina Adenina HO-CH2 0 HO-CH2 H H H H H H CH O. -0- CH .0. -CHO. H H H H HI HI II H H A H H OH OH OH OHI OH OH OH OH Adenosina Guanosina Timidina Citidina Adenina 0 HO-CHg 0 Adenina H H 5 H O-P- O-CH H H 1 OH H H O P-O OH OH NII NH. HIN N HN IIN O 0 ÓH 3 Adenosina 3'-monofosfato Adenosina 2',3'-monofosfato cíclico Estos mononucleótidos pueden incorporar el fosfato en posiciones diferentes al carbono 5' del azúcar, pueden unir más de un fosfato, denominándose di y trinucleótidos e incluso pueden tener los grupos fosfatos unidos a más de un carbono y ser cíclicos.
H H H H -0-CH2_0. 2 0 Desoxicitidilato3
Enlace Fosfodiéster y Polinucleótidos
Los nucleotidos se unen entre sí para formar la molecula de DNA y RNA mediante un enlace denominado fosfodiester. En este tipo de enlace el grupo hidroxilo del carbono 3' reacciona con el grupo hidroxilo del carbono 5' del siguiente nucleótido con la participación de un grupo fosfato. Generalmente un ácido nucleico corto se denomina oligonucleótido, y aquellos de más de 50 nucleótidos se denominan polinucleótidos.
El DNA (Ácido Desoxirribonucleico)
El DNA es un polímero lineal de residuos desoxirribonucleicos unidos por enlace fosfodiéster entre las posiciones 3' y 5' de las moléculas de 2'-desoxiribosa. Las bases del DNA son adenina, guanina, timina y citosina. La diferencia entre dos moléculas de DNA reside en la secuencia de bases. Esta secuencia lineal de residuos es lo que forma la estructura primaria del DNA. Un extremo de la cadena presenta libre un grupo 5'-OH y el otro un grupo 3'-OH y por acuerdo cuando se nombra una cadena, se hace tomando en primer lugar la base correspondiente al extremo 5'. El tamaño de las moléculas varía desde unos cientos de residuos en algunos virus hasta 109 residuos en el DNA cromosómico humano.
ADN ARN Para la formulación definitiva del modelo estructural del DNA fueron necesarias las contribuciones de varios científicos. En 1940 Chargaff estudiando el DNA procedente de un gran numero de distintas especies concluye:
- La composición de bases del DNA varía de una especie a otra.
- Las muestras de tejidos diferentes de una misma especie tienen la misma composición de bases.
- La composición de bases de una especie, no varia con la edad, nutrición ni condiciones ambientales.
- En todas las especies la cantidad de adeninas es igual a la de timinas y la de citosina a la de guanina, o lo que es lo mismo la suma de purinas (A y G) es igual a la de pirimidinas (C y T).
Extremo 5' = FCH2_0 =CH2 0 H H Enlace O OH fosfodiéster 0-P-0 T C & CH_ 0 H H H FH H ÓH -P=0 0-P=0 C € C ICH Q H H H H CH H OH Op 0-P 0 C 0 C "CH,0 H H H = H 0 0-P-0 0-P-0 O A - H H H OH Extremo 3' Rosalind Franklin y M. Wilkins aportaron mediante el estudio con técnicas de difracción de rayos X que las moléculas de DNA eran helicoidales con dos periocidades a 3,4 Å y 34 Å y para finalizar, con todos los datos anteriores, la estructura que adoptan las fibras de DNA (o estructura secundaria) fue deducida en 1953 por Watson y Crick. El modelo estructural que propusieron se resumiría en los siguientes puntos:
- Las moléculas de DNA se componen de dos cadenas antiparalelas helicoidales enrolladas alrededor de un mismo eje, formando una doble hélice dextrógira.
- Cada purina de una cadena de DNA de doble hélice se une a una pirimidina de la cadena complementaria, así, la A se une a la T por dos puentes de hidrógeno y la G a la C por tres. Las moléculas de pentosa y el grupo fosfato quedan hacía afuera y las bases nitrogenadas, que son hidrofóbicas y prácticamente planas están apiladas en el interior y en posición perpendicular al eje. La estructura se estabiliza por tanto, por puentes de hidrógeno entre bases complementarias y por las interacciones de apilamiento de bases.
A CH2 0 H C4
- El diámetro de las hélices es de 20 Å (2 nm), las bases adyacentes están separadas por 3.4 Å y rotan 36º, por tanto la doble hélice completa una vuelta cada diez residuos o lo que es igual cada 34 Å. En realidad, en disoluciones acuosas se ha demostrado que el DNA-B tiene 10,5 residuos por vuelta, lo que es igual a unos 36 Å.
- La conformación espacial de las dos cadenas da lugar en la superficie de la hélice a la formación de un surco mayor y un surco menor.
Este modelo propuesto por Watson y Crick es el modelo conocido como DNA-B y es la estructura más estable y por tanto frecuente, que adopta el DNA en condiciones fisiológicas. Otros modelos estructurales son el DNA-A que es la forma que adquiere el DNA-B cuando se deshidrata o en soluciones con poco agua; la hélice es más gruesa, el surco mayor más profundo, el menor más superficial y el numero de pares de bases por vuelta es once. Debido a los recativos empleados, los DNAs cortos cristalizan en esta forma. El DNA-Z difiere del B principalmente por ser una molécula levógira, contiene doce pares de bases por vuelta y tiene una estructura más delgada y alargada, el surco mayor es apenas observable y el menor estrecho y profundo. La presencia de DNA-A en las células no se ha podido confirmar hasta el momento, sin embargo, pequeñas zonas o fragmentos de DNA-Z se encuentran en células procariotas y eucariotas. Esta estructura del DNA, se asocia con la regulación de la expresión de algunos genes o con la recombinación genética.
# *- 28 Å CH 5 3' H H WW H ~8.0 A* NIN H-N Thymine N H- C-1 ! Adenine C N H C-1 -11.1 Â *- 29A H ₭~3.0 À~1 OZ Cytosine Guanine C-1" T H ~29AX 28 Å -10.8 A- 3' 5 La disposición de las distintas moléculas de DNA de los distintos individuos (circular, lineal, alternativo) da lugar a una estructura terciaria denominada superenrollamiento. El DNA dúplex está enrollado a la derecha, si el torcimiento de la doble hélice alrededor de su propio eje es también a la derecha, el DNA se superenrolla positivamente, y si lo hace en sentido contrario, negativamente. Este superenrollamiento se observa en el DNA lineal de plantas y animales, así como en el DNA circular de bacterias.
Estructuras no Habituales en las Secuencias de DNA
Se trata de secuencias curiosas estructuralmente que podemos encontrarnos en el DNA y que con certeza tendrán alguna consecuencia funcional, como ejemplo tenemos que la acumulación de adeninas consecutivas en una hebra produce la curvatura de la hélice.
" H N-HIN
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
