Le biosintesi dei lipidi e il metabolismo del colesterolo
Slide di Università sulle biosintesi dei lipidi e il metabolismo del colesterolo. Il Pdf, adatto per lo studio universitario di Biologia, illustra i processi di digestione e assorbimento dei lipidi, il ruolo dei sali biliari e l'aterosclerosi.
Mostra di più36 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
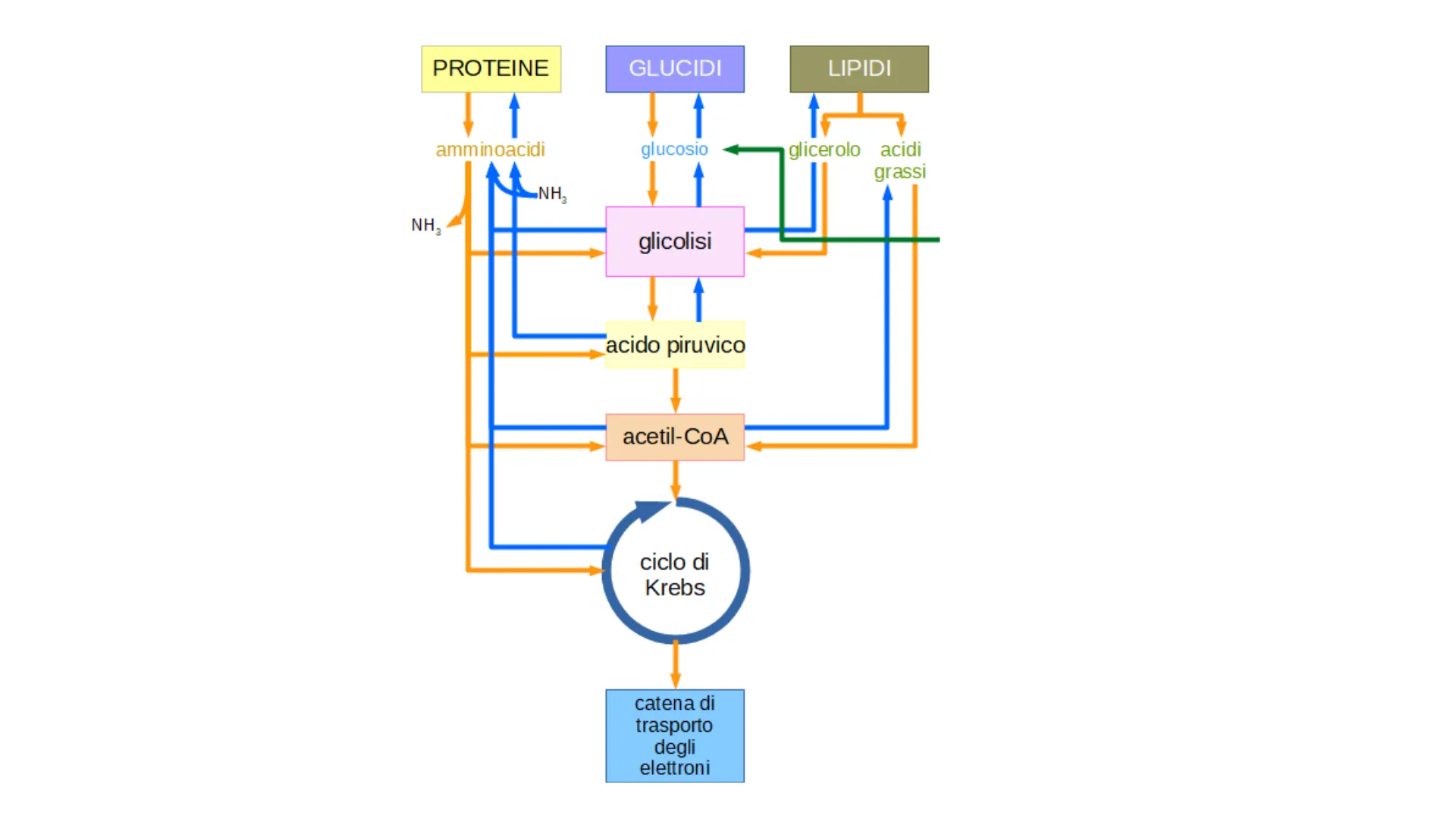
Le biosintesi dei lipidi
PROTEINE GLUCIDI LIPIDI amminoacidi glucosio glicerolo acidi grassi NH2 NH glicolisi acido piruvico acetil-CoA ciclo di Krebs catena di trasporto degli elettroni
- I triacilgliceroli (TAG) sono la principale fonte di accumulo di energia (tessuto adiposo)
- Sono degradati quando serve energia
- Sono sintetizzati a partire da glucosio e amminoacidi quando ci sia un eccesso di questi substrati energetici
- La sintesi avviene nel fegato e nelle cellule epiteliali mammarie durante la lattazione - scarsa nel tessuto adiposo
- La sintesi ha inizio da acetil CoA
- Via degradativa e via sintetica hanno localizzazione diversa e sistemi enzimatici differenti - regolate in modo reciproco
Beta ossidazione è mitocondriale Sintesi degli acidi grassi è citoplasmatica
Trasferimento di acetil CoA nel citoplasma
Prima tappa della biosintesi degli acidi grassi: trasferimento di acetil CoA nel citoplasma
- Acetil CoA derivato dal glucosio e amminoacidi si accumula nella matrice mitocondriale
- Se in eccesso è trasferito nel citoplasma per la sintesi degli acidi grassi
- Il trasferimento avviene sotto forma di citrato
- E' stimolato dall'aumento della concentrazione del citrato stesso nella matrice mitocondriale: segnala che c'è un eccesso di energia.
NB: ricordare che il citrato deriva dalla prima reazione del ciclo di Krebs (ossalacetato + acetil CoA) MATRICE MITOCONDRIALE Ossalacetato (OAA) Acetil CoA H2O Citrato sintasi CoA Citrato MEMBRANA MITOCONDRIALE INTERNA Citrato CoA. ATP H2O- ATP-citrato liasi ADP + Pi OAA Acetil CoA CITOSOL
Si forma È utilizzato per la sintesi MITOCHONDRION CYTOPLASM Acetyl COA Acetyl COA Citrate >Citrate Oxaloacetate NADH Oxaloacetate Malate 1 Pyruvate Pyruvate NADPH
Il sistema navetta di trasporto del citrato è piuttosto complesso nell'insieme assicura anche il rientro dell'ossalacetato sotto forma di malato e produce NADPH
Formazione del malonil CoA
La formazione del primo intermedio della sintesi: il malonil CoA
Malonil Co-A è il primo intermedio della sintesi degli acidi grassi Sintetizzato da Acetil CoA carbossilasi Utilizza la vitamina biotina (Vit H o B8) e consuma ATP, CO2 La reazione è limitante per la velocità della sintesi ed il punto di regolazione Regolazione allosterica: stimolata da citrato (= energia abbondante) e inibita da eccessi di acil-CoA a lunga catena
Catena laterale H Braccio della Lys della biotina H 0= S Proteina trasportatrice della biotina 0 HCO + ATP +ADP + P. -NH O -c=0 HN Biotina carbossilasi Transcarbossilasi Proteina trasportatrice della biotina Acetil-CoA HC-C. =0 S-CoA 0= NH 180° Biotina carbossilasi Transcarbossilasi Proteina trasportatrice della biotina 0 HN + -CH .- VH 0. S-CoA Malonil-CoA NH Proteina trasportatrice della biotina
Acetil CoA carbossilasi (protomeri inattivi) Citrato + KUIIII Acil CoA grasso a lunga catena Acetil CoA carbossilasi (polimero attivo) C O O CH3-C-S-CoA C-CH2-C-S-CoA Acetil CoA O Malonil CoA CO2 ATP ADP + P
Regolazione dell'acetil CoA carbossilasi
Regolazione ormonale ed a lungo termine dell'acetil CoA carbossilasi
Regolazione ormonale della acetil CoA carbossilasi: Glucagone e adrenalina la inattivano fosforilandola Si verifica quando glicemia è bassa e quando acetil CoA va usato per produrre energia Insulina la attiva defosforilandola Si verifica quando glicemia è alta e si possono accumulare acidi grassi. NB: avviene l'inverso per la regolazione della beta-ossidazione che degrada gli acidi grassi Regolazione a livello genico: diete abbondanti stimolano la sintesi dell'enzima, diete ipocaloriche tendono a ridurla
+ Insulina Proteina fosfatasi Pi 1 Acetil CoA carbossilasi (inattiva) P Acetil CoA carbossilasi (attiva) + _ Proteina chinasi attivata dall'AMP (AMPK) ADP ATP + AMPK chinasi (covalente) AMP (allosterico) + Glucagone Adrenalina (covalente)
Sintesi degli acidi grassi: acido grasso sintasi
La sintesi degli acidi grassi: l'acido grasso sintasi
La acido grasso sintasi dei mammiferi (FAS I) è un grosso polipeptide con 7 diversi domini strutturali che svolgono funzioni diverse. Sintetizza catene di acido palmitico (16:0 - 16 atomi di C, nessun doppio legame) e poi le rilascia In pratica ha la funzione di enzimi multipli che sono all'interno di una stessa lunga catena polipeptidica In particolare un dominio della proteina - la proteina trasportatrice di acili - mantiene la catena in crescita legata alla proteina e la sposta sui vari siti attivi che la modificano
Il processo ripete una serie di quattro passaggi che ogni volta allungano la catena di 2 atomi di carbonio In due dei passaggi ripetuti della sintesi serve NADPH come coenzima ridotto donatore di protoni ed elettroni.
CH3 CH CH 1 CH2 - CH, CH2 - CH2 CH2 1 CH2 CH 1 CH2 CH2 1 CH2 Figura 21.3 La sintesi di palmitato. La catena dell'acido grasso cresce di due atomi di carbonio per volta, donati dal malonato attivato, con perdita di una molecola di CO2. Il gruppo acetilico iniziale è ombreggiato in giallo; il C-1 e il C-2 del malonato sono ombreggiati in rosa, e il carbonio rilasciato sotto forma di CO2 è in verde. Dopo l'aggiunta di ogni unità bicarboniosa, due reazioni di riduzione convertono la catena nascente in acido grasso saturo con quattro, sei, otto e così via atomi di carbonio. Il prodotto finale è il palmitato (16:0).
HS HS CH 1 C 0 Palmitato CH2-COO CH3 CH3 CH3 CH2-COO C=0 CH2 1 CH, CH, CH3 C=0 CH2 CH2 C=0 S CH2-COO CHO CH, S 4H+ C=0 CH, S CH2-COO- CH2 C=0 CH2 + - S C=0 1 S altre CO2 quattro addizioni Acido grasso sintasi 5 1 CO2 CO. 1 S F 4H+ + S 4H+ CH2 4e + 4 4e C=0 C=0 S CH, + 4eCH2-COO- CH3 - CH2-COO- CH2 C=0 CH2 1 CH3 S CH2 CH2 1 CH2- COO CH2 S 4H+ C=0 4H+ CH2 CH2 - CH2-COO- 1 C=0 4H+ CH2 1 S 4e C=0 4 - S 1 altre quattro addizioni CH, CH2 CH2 HȘ CH2 HS CH2 + -0-0 O Palmitato CH3 CH2 CH2 CH2 CH2 CH2 CH2 1 CH2 CH2 CH2 Figura 21.3 La sintesi di palmitato. La catena dell'acido grasso cresce di due atomi di carbonio per volta, donati dal malonato attivato, con perdita di una molecola di CO2. Il gruppo acetilico iniziale è ombreggiato in giallo; il C-1 e il C-2 del malonato sono ombreggiati in rosa, e il carbonio rilasciato sotto forma di CO2 è in verde. Dopo l'aggiunta di ogni unità bicarboniosa, due reazioni di riduzione convertono la catena nascente in acido grasso saturo con quattro, sei, otto e così via atomi di carbonio. Il prodotto finale è il palmitato (16:0). CH3 CH3 CH2 C=0 CH2 C=0 S C=0 CH2 + S n 4e 1 + S 4 4e C=0 CH2 1 1 4 - S 4 + CO2 CO2 CO2 Acido grasso sintasi 5
Dopo 7 cicli di condensazione si produce una molecola di acido palmitico La reazione consuma ATP e NADPH
KS MT ACP AT 1 KR ER HD (ER) Enoil-ACP reduttasi (KR) ß-chetoacil-ACP reduttasi ER KR (DH) ß- idrossiacil-ACP deidratasi DH ACP (ACP) proteina trasportatrice di acili HS SH KS MAT (KS) ß-chetoacil -ACP sintasi (MAT) Malonil/acetil-CoA-ACP transferasi
8 acetil CoA + 7 ATP + 14 NADPH + 14 H+ -palmitato + 8 CoA + 7 ADP + 7 Pi + 14 NADP+ + 6 H2O
- Il NADPH necessario è abbondante nei tessuti in cui la sintesi avviene (fegato, ghiandola mammaria)
Produzione di NADPH
Il NADPH necessario è abbondante nei tessuti in cui avviene la sintesi degli acidi grassi (fegato, ghiandola mammaria) E' prodotto da una via metabolica specializzata: la via dei pentoso fosfato
NADP+ NADPH NADP+ NADPH Glucose 6-phosphate 1 Ribulose 5-phosphate pentose phosphate pathway
Anche l'enzima malico permette di produrre NADPH partendo dal malato.
COO- NADP+ NADPH + H+ COO- - CHOH 1 C=0 CH2 malic enzyme CH3 COO- Malate Pyruvate I + CO2
Via del pentosio fosfato
Via del pentosio fosfato
Importante in fegato, ghiandola mammaria, t. adiposo Non serve a produrre energia
- Produce NADPH, forma ridotta del coenzima che è usato nelle biosintesi dei lipidi
- Produce ribosio 5P usato nella sintesi dei nucleotidi
- Converte pentosi in intermedi metabolici della glicolisi.
NADPH è utilizzato per proteggere dai processi ossidativi generati dal metabolismo cellulare (respirazione mitocondriale)
Fase non ossidativa Fase ossidativa Glucosio 6-fosfato NADP+ 2 GSH glutatione reduttasi NADPH GSSG transchetolasi, transaldolasi 6-Fosfogluconato Acidi grassi, steroli, ecc. NADP+< biosintesi riduttiva CO2 NADPH Precursori Ribulosio 5-fosfato Ribosio 5-fosfato Nucleotidi, coenzimi, DNA, RNA Nelson & Cox I principi di Biochimica Zanichelli 6 ed.
Modificazioni degli acidi grassi
Modificazioni degli acidi grassi: allungamento della catena e sintesi di grassi polinsaturi
Attraverso vie metaboliche specializzate l'acido palmitico può essere allungato (+ 2C) a formare acidi grassi saturi più lunghi L'introduzione di un doppio legame in posizione 49 permette la sintesi di acidi grassi insaturi/polinsaturi La capacità delle cellule animali di sintetizzare acidi grassi insaturi è limitata: i grassi omega 3 e omega 6 non possono essere sintetizzati Gli omega 3 e omega 6 sono acidi grassi essenziali Hanno ruoli biologici di estrema importanza
Palmitato 16:0 desaturazione allungamento Palmitoleato 16:1 (49) Stearato 18:0 allungamento desaturazione Acidi grassi saturi più lunghi Oleato 18:1 (49) desaturazione (solo nelle piante) Omega 6 Linoleato 18:2 (49,12) desaturazione (solo nelle piante) desaturazione Omega 3 «-Linolenato 18:3 (49,12,15) y-Linolenato 18:3 (A6,9,12) allungamento Acido eicosapentanoico (EPA; 20:5 (45,8,11,14,17)) Eicosatrienoato 20:3 (48,11,14) desaturazione Acido docosaesanoico (DHA; 22:6(4 4,7,10,13,16,19)) Arachidonato 20:4 (45,8,11,14)
Funzioni degli acidi grassi polinsaturi
Gli acidi grassi polinsaturi sono importanti in diverse funzioni biologiche -fluidità delle membrane -precursori di molecole bioattive (prostaglandine, trombossani, leucotrieni) -regolatori del processo infiammatorio
UNSATURATED FATTY ACID H H H H H H H C-C-c-c-c-C- H C H 0 1 H H H H OMEGA END 1 H -I -H H SATURATED FATTY ACID OS | OMEGA END C-C-C- -C-C- C- C-C-C-H 1 O H I-U-I H I-U-I I-U-I I- U-I I-U-I H H -I H 1 H @ Byjus.com linoleic acid (9,12-octadecadienoic) a 0 O 9 12 6 omega-6 fatty acid linolenic acid (9,12,15-octadecadienoic) a 0 9 12 15 O 3 omega-3 fatty acid H H H 1 1 1 CE 1 H -C-C 1 O
Sintesi dei triacilgliceroli e fosfolipidi
La sintesi dei triacil gliceroli e dei fosfolipidi
Destino degli acidi grassi: 1. essere incorporati nei triacilgliceroli (TAG) 2. Essere utilizzati per la sintesi dei glicerofosfolipidi - componenti della membrana cellulare I TAG sono la più importante forma di riserva di energia nei mammiferi, assicurando un apporto energetico (teorico) della durata di settimane
- In entrambi i casi il primo passaggio è la sintesi di glicerolo 3 fosfato 1. da diidrossiacetone fosfato 2. da glicerolo
Glucosio 4 glicolisi HCOH 1 C=0 0 1 HC-0-P-O" HCOH 1 HCOH 1 H_COH 1 Diidrossiacetone fosfato Glicerolo NADH + H+ ATP glicerolo 3-fosfato deidrogenasi glicerolo chinasi ADP NAD+ H COH HO-C-H O I HC-0-P-O" 1 0 I .-- Glicerolo 3-fosfato
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
