Trasporti passivi: diffusione facilitata, canali ionici e trasportatori
Documento sui trasporti passivi: diffusione facilitata. Il Pdf esplora i meccanismi di passaggio delle molecole attraverso le membrane cellulari, la selettività e la regolazione di questi processi, con un focus sui trasportatori GLUT del glucosio, utile per lo studio della Biologia a livello universitario.
Mostra di più13 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
TRASPORTI PASSIVI: DIFFUSIONE FACILITATA
Abbiamo visto che la diffusione semplice consente il passaggio di molecole attraverso le membrane biologiche solo di un ristretto gruppo di molecole, per lo più molecole polari, piccole polari, molecole idrofobiche e gas.
Esistono però una serie di molecole escluse dalla diffusione semplice, ma d'altro canto sono comunque molecole di estrema importanza per la sopravvivenza della cellula:
- Grosse molecole polari, come glucosio e amminoacidi
- Molecole cariche, gli ioni.
Come riescono ad attraversare il doppio strato lipidico queste molecole, considerando che la loro natura non consente loro di attraversa il core lipidico in modo diretto?
- Diffusione facilitata
Non richiede altra energia che quella del moto molecolare
Trasporti passivi
- Non richiedono energia
- Vanno da concentrazione maggiore a quella minore
- Sono equilibrativi
Diffusione Diffusione semplice Diffusione facilitata Canali ionici Trasportatori
Il passaggio di queste molecole, che altrimenti non potrebbe avvenire all'interno del core lipidico del doppio strato, è reso possibile dalla presenza all'intento della membrana plasmatica di strutture proteiche:
- Canali ionici
- Trasportatori
Che creano un poro acquoso all'interno del quale le grosse molecole organiche/polari/cariche, possono attraversare il doppio strato lipidico. Si tratta ancora di un trasporto passivo, per cui: · Non richiede energia, se non quella del moto molecolare · Va sempre secondo gradiente, quindi da dove la C è maggiore a dove è minore · È anch'esso equilibrativo, cioè il trasporto continua fintanto che la C nello spazio extra e intracellulare raggiunge lo stesso valore.
Canali ionici Trasportatori o carrier LEC Membrana cellulare LIC Trasportatore aperto verso il LIC Stesso trasportatore aperto verso il LEC
Le proteine che rendono possibile il passaggio di queste molecole attraverso il doppio strato lipidico, sono grossolanamente raggruppate in due categorie:
Canali ionici: sono proteine che formano dei veri e propri canali, sono normalmente proteine a più domini transmembrana che si associano per formare un canale intendo acquoso, che rende possibile il passaggio di molecole; normalmente consentono il passaggio di ioni e acqua. I canali ionici sono contemporaneamente accessibili dal lato extracellulare e da quello intracellulare.
Trasportatori o carrier: sono anch'esse delle proteine integrali di membrana e dal punto di vista strutturale, così come i canali ionici, sono formate soprattutto da proteine polimorfiche. Anche in questo caso si viene a formare un core idrofilo all'interno del quale possono passare le molecole, che sono fondamentalmente molecole organiche, ma anche ioni: la grossa differenza dai canali ionici è che queste proteine non sono contemporaneamente accessibili dal lato extra e intracellulare, ma sono alternativamente accessibili ai due domini. Questo perchè normalmente, nelle proteine trasportatrici o carrier, esiste un'interazione diretta tra la molecola che deve essere trasportata e il trasportatore stesso. Cioè esiste una sorta di legame ad affinità variabile ed è proprio questo legame con la molecola che deve essere trasportata, che dà origine a questi cambiamenti conformazionali, che rende la molecola alternativamente accessibile da una parte o dall'altra della membrana plasmatica.
Gas Molecole idrofobiche Piccole Grosse Molecole cariche H H,O Ca2+ CO, Na+ Ioni O2 Benzene Etanolo Glucosio Amminoacidi molecole polari molecole polari
Diffusione facilitata - Canali
Channel through center of membrane protein viewed from above Channel through center of membrane protein
Le proteine che formano i canali ionici sono a più domini transmembrana: ciascuna alfa-elica di queste proteine normalmente si dispone all'intento del doppio strato creando un poro centrale. Di solito i canali ionici sono delle proteine a più subunità e le varie subunità formano proprio la struttura centrale idrofila. Questo poro all'interno della proteina è una via di passaggio permeabile e contemporaneamente accessibile dal lato extracellulare e intracellulare.
CANALI DEL K+VOLTAGGIO DIPENDENTI
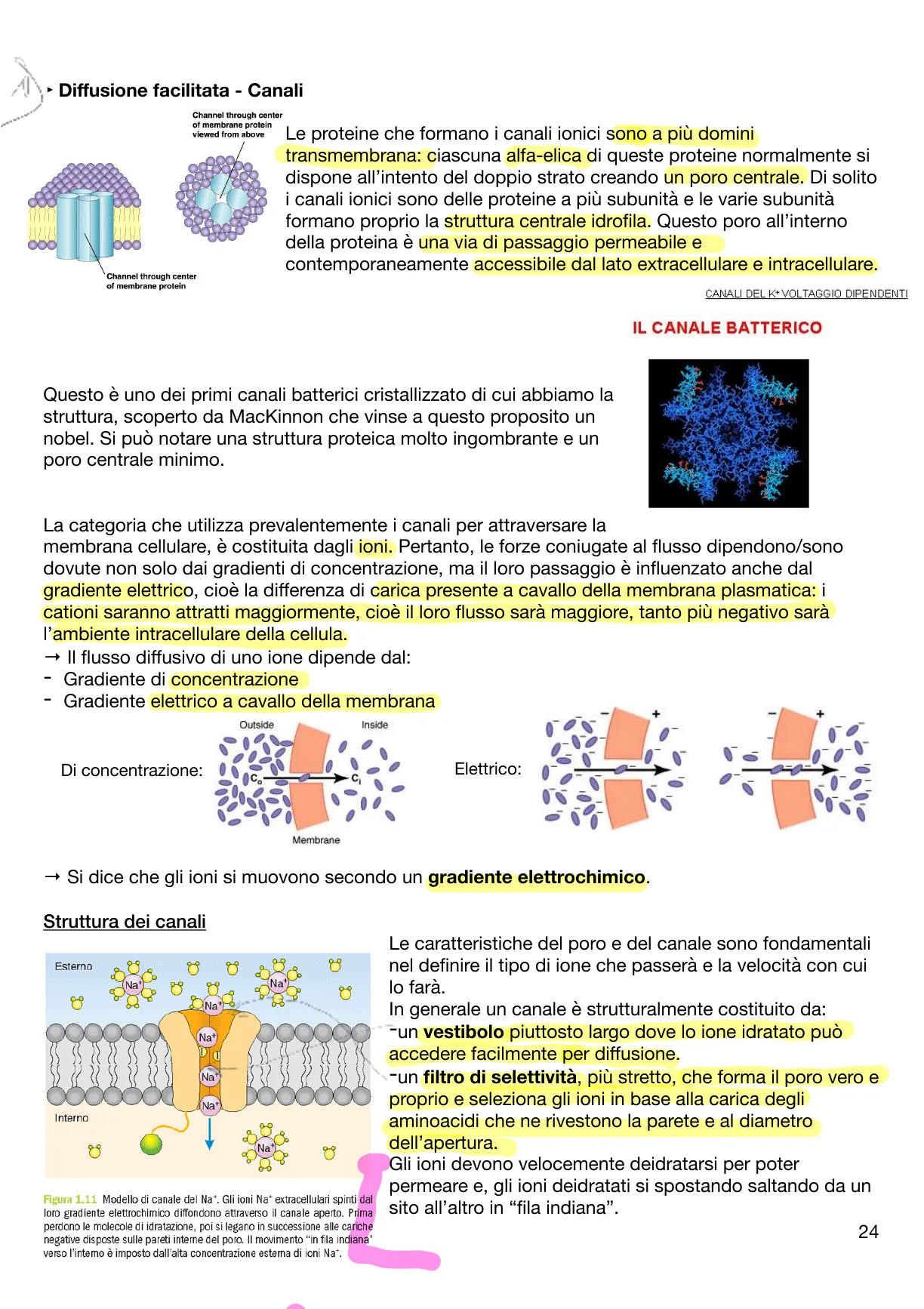
IL CANALE BATTERICO
Questo è uno dei primi canali batterici cristallizzato di cui abbiamo la struttura, scoperto da Mackinnon che vinse a questo proposito un nobel. Si può notare una struttura proteica molto ingombrante e un poro centrale minimo.
La categoria che utilizza prevalentemente i canali per attraversare la membrana cellulare, è costituita dagli ioni. Pertanto, le forze coniugate al flusso dipendono/sono dovute non solo dai gradienti di concentrazione, ma il loro passaggio è influenzato anche dal gradiente elettrico, cioè la differenza di carica presente a cavallo della membrana plasmatica: i cationi saranno attratti maggiormente, cioè il loro flusso sarà maggiore, tanto più negativo sarà l'ambiente intracellulare della cellula.
- Il flusso diffusivo di uno ione dipende dal:
- Gradiente di concentrazione
- Gradiente elettrico a cavallo della membrana
+ + Outside Inside Di concentrazione: C. Elettrico: Membrane
- Si dice che gli ioni si muovono secondo un gradiente elettrochimico,
Struttura dei canali
Esterno Na+ Na+ Na+ Na+ D Na+ Na+ Interno Na
Figura 1.11 Modello di canale del Na+. Gli ioni Na+ extracellulari spinti dal loro gradiente elettrochimico diffondono attraverso il canale aperto. Prima perdono le molecole di idratazione, poi si legano in successione alle cariche negative disposte sulle pareti interne del poro. Il movimento "in fila indiana' verso l'interno è imposto dall'alta concentrazione esterna di ioni Na+.
Le caratteristiche del poro e del canale sono fondamentali nel definire il tipo di ione che passerà e la velocità con cui lo farà.
In generale un canale è strutturalmente costituito da:
- un vestibolo piuttosto largo dove lo ione idratato può accedere facilmente per diffusione.
- un filtro di selettività, più stretto, che forma il poro vero e proprio e seleziona gli ioni in base alla carica degli aminoacidi che ne rivestono la parete e al diametro dell'apertura.
Gli ioni devono velocemente deidratarsi per poter permeare e, gli ioni deidratati si spostando saltando da un sito all'altro in "fila indiana".
24Dal punto di vista funzionale i canali si distinguono in base a due caratteristiche:
1. Selettività
Si basa sulla capacità che ha la struttura a canale di selezionare uno ione. Normalmente abbiamo due categorie di canali ionici:
- Canali con alta selettività, quindi fanno passare un solo ione:
- del Na+
- del K+
- del Ca2+
- del CI-
- Canali con selettività intermedia, che lasciano passare più categorie di ioni:
- del Na+, K+, Ca2+ (cationici)
- Anionici
- Del Na+ e K+ (cationici monovalenti)
filtro di selettività = I camera interna 'sensore" del voltaggio "gate" di attivazione "gate" di inattivazione
2. Modalità di attivazione
All'interno della membrana plasmatica ci sono due tipi di canali:
- Canali aperti: sono accessibili in ogni momento e ad entrambi i lati della membrana, come ad esempio l'acquaporina o alcuni canali ionici; il numero di questi canali è molto limitano, altrimenti avremmo un passaggio continuo di cariche dall'interno all'esterno della membrana. Nella maggior parte dei casi sono presenti i cosiddetti canali a cancello.
CANALI APERTI CANALI A CANCELLO LEC LEC Aperto Aperto Chiuso
- Canali a cancello: sono così detti perchè normalmente chiusi e quando i messaggeri secondari, o molecole disponibili a livello extra o intracellulare, si legano al canale, ne determinano l'apertura.
Voltaggio- dipendente Ligando- dipendente (extracell) Ligando- dipendente (intracell) Meccano- dipendente Temperatura- dipendente
I canali a cancello sono normalmente chiusi. Si aprono in risposta a segnali: -Elettrici (potenziale di membrana) chiuso -Chimici (Ligandi intracellulari o estracellulari) -Meccanici -Termici ! ! aperto CYTOSOL extracellulare
Saturazione parziale
Diffusione attraverso canali ionici
Figura 1.10 Relazione tra permeabilità (velocità di trasporto attraverso la membrana) e la concentrazione di soluti nel liquido extracellulare. Nel caso della diffusione passiva la relazione è lineare. Nel caso della diffusione faci- litata (carrier) o attraverso canali ionici, la relazione è invece lineare a basse concentrazioni per poi saturare in maniera diversa ad alte concentrazioni.
Dal punto di vista cinetico, il trasporto passivo mediante diffusione facilitata operata da canali ionici, ricorda molto la diffusione semplice.
Se mettiamo in ordinate la velocità di trasporto in funzione della concentrazione dello ione nell'ambiente di partenza, abbiamo che per concentrazioni medio-alte, ma comunque in generale nel range fisiologico, all'aumentare della concentrazione dello ione nell'ambiente di partenza aumenta anche la velocità di trasporto.
Per concentrazioni estremamente elevate, che si incontrano raramente nei sistemi biologici, si h una curva che tende a saturare perchè a questo punto l'elemento limitante diventa il numero di canali ionici presenti all'interno della membrana plasmatica.
Molecole - ioni, acqua Forza motrice - gradiente elettrochimico
Lezione 16/10 Diffusione facilitata - Trasportatori
Extracellular fluid Intracellular fluid Molecule to be transported Gate to intracellular fluid closed
Anche queste proteine sono costituite da più subunità e formano un poro acquoso. La grande differenza è data dal fatto che queste proteine normalmente non sono Carrier contemporaneamente accessibili dal lato extra e intracellulare. Pertanto, essi esistono in almeno tre stati conformazionali:
Membrane
- Proteina aperta verso il lato extracellulare: quindi è accessibile alle molecole presenti da questo lato.
- Proteina apparentemente chiusa (stato intermedio): la molecola che deve essere trasportata si trova all'interno di questo poro acquoso.
- Proteina aperta verso il lato intracellulare: la molecola trasportata può essere qui rilasciata.
Mentre nei canali ionici gli ioni passavano in fila indiana, nei trasportatori le molecole che devono essere trasportate sono molecole che si legano selettivamente alla proteina carrier.
Le proteine carrier si legano a substrati specifici e li trasportano attraverso la membrana cambiando conformazione.
- Uniporti = sono in grado di trasportare una sola molecola alla volta.
- Substrati = piccole molecole organiche: amino acidi, glucosio, nucleotidi, metaboliti.
- Il trasporto procede secondo gradiente, in entrambe le direzioni.
Come cambia conformazione il trasportatore?
000. 0 000 000
Normalmente i domini transmembrana di queste proteine trasportatrici sono alfa-eliche, che si dispongono nel doppio strato lipidico a formare queste strutture molto orientate. Grazie all'orientamento di queste alfa-eliche la struttura centrale del carrier, il poro, è accessibile dal lato extracellulare, ma non dal lato intracellulare. La molecola di glucosio presente in grande concentrazione nell'ambiente extra trova un sito di legame specifico all'interno di questa struttura a doppia elica. Il legame della molecola di glucosio con gli amminoacidi presenti all'interno della struttura a doppia elica, determina un cambiamento conformazionale: semplicemente è un ribaltamento/spostamento di pochi gradi di queste alfa-eliche, sufficiente per rigenerare questo stato conformazionale differente
000 000 000 000 000
È semplicemente cambiata l'angolatura con cui si incrociano queste alfa- eliche, per generare una struttura che questa volta è chiusa sul lato extracellulare e aperta sul lato intracellulare. La molecola si stacca e tale distacco genera nuovamente il cambiamento conformazione che ribalta la proteina e la ridispone ad un nuovo ciclo di trasporto.
26 Gate to extracellular fluid closed
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
