Biochimica d'organo: metabolismo glutammato e GABA nel sistema nervoso
Documento dall'Università degli Studi di Torino su Biochimica: Modulo di Biochimica D'organo. Il Pdf, un approfondimento di Biologia a livello universitario, esplora il sistema nervoso centrale, il metabolismo del glutammato e del GABA, e i meccanismi di vasocostrizione e vasodilatazione cerebrale.
Mostra di più20 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
LEZIONE 12 - BIOCHIMICA D'ORGANO
Introduzione al Corso
La prof.ssa avvisa che sarà necessario un cambio orario nella settimana del 20 maggio e, avendo anticipato l'inizio delle sue lezioni, in quella settimana ci sarà una lezione in meno, a meno che si sfruttino le lezioni del 21/22 maggio e si scalino dalla fine.
Per le lezioni opzionali con il professor Stern è già stato programmato il calendario. Se ci fossero cambi è possibile comunicarli qualche giorno prima delle lezioni.
Per contattare la prof.ssa si può utilizzare la mail: chiara.riganti@unito.it
Il materiale didattico è reperibile sulla piattaforma moodle nel corso di biochimica d'organo: https://elearning.unito.it/medicina/course/view.php?id=14909 (BIOCHIMICA D'ORGANO_BIOCHIMICA_TORINO_CBGO_SME0980C_Anno_1_24_25)
Testi Consigliati
I libri consigliati sono gli stessi presentati all'inizio del corso di biochimica metabolica, perché un libro solo per biochimica d'organo non esiste (alcuni testi hanno delle parti più approfondite e altri meno), quindi basta prendere le informazioni da un eventuale testo acquistato ad inizio anno. In linea di massima, però, ciò che c'è nelle diapositive, è tutto quello che può essere argomento d'esame.
- Nelson-Cox. Lehninger principles of biochemistry. Zanichelli
- Devlin. Textbook of biochemistry. EdiSES
- Berg, Tymoczko, Stryer. Biochemistry. Zanichelli
- Mathews, van Holde, Appling, Cahill. Biochemistry. Piccin
- Garrett, Grisham. Biochimica. Piccin
Programma del Corso
Il programma inizia con il tessuto cerebrale, per poi analizzare il sistema intestinale, sistema endocrino, rene, ormoni, tessuto scheletrico, tessuto connettivo. Alcuni argomenti, trattando contemporaneamente i corsi di istologia, anatomia e biochimica, sono collegati tra di loro.
- Biochimica del sistema nervoso centrale: metabolismo in condizioni fisiologiche delle cellule neuronali e gliali. Alterazioni del metabolismo in condizioni di ipossia. Basi biochimiche delle patologie neurodegenerative da alterato folding proteico.
- Biochimica del tratto gastrointestinale: composizione dei fluidi gastrointestinali; sistema neuro-endocrino intestinale; sistemi di secrezione e assorbimento del tratto gastrointestinale; funzioni del pancreas e del fegato basi biochimiche di alcune patologie che affliggono il tratto gastrointestinale: morbo celiaco, malattie infiammatorie intestinali, fibrosi cistica.
- Biochimica del fegato: vie metaboliche peculiari del fegato; funzione digestiva del fegato: sintesi e catabolismo degli acidi biliari; funzioni di detossificazione di cataboliti endogeni (bilirubina) e xenobiotici.
- Biochimica del sistema endocrino: sintesi ed effetti dei seguenti ormoni: ormone somatotropo; ormoni tiroidei; glucocorticoidi surrenalici; androgeni; estrogeni e progesterone; ossitocina; prolattina.
- Biochimica del rene: sistemi di trasporto e riassorbimento. Omeostasi del Na+ e del K+. Equilibrio acido base. Bilancio idrico. Sintesi ed effetti degli ormoni coinvolti in tali processi: sistema renina-angiotensina-aldosterone; ormone antidiuretico; peptide natriuretico atriale.
- Biochimica del tessuto scheletrico. Omeostasi del calcio e del fosfato. Sintesi ed effetti di vitamina D, paratormone e calcitonina.
- Biochimica del tessuto connettivo: composizione e struttura del tessuto connettivo: sintesi e funzioni di glicosamminoglicani, collagene, elastina. Basi biochimiche di alcune connettivopatie: collagenopatie, condrodisplasie, sindromi carenziali (scorbuto).
Se ci fosse necessità di seguire le lezioni a distanza la prof.ssa è disponibile a creare lezioni su Webex: https://unito.webex.com/meet/chiara.rigant.
La prof.ssa si dimostra molto disponibile per chiarire qualsiasi dubbio riguardo alla sua parte di programma, sia metabolica sia d'organo.
ESAME
Il primo appello è a fine giugno e il secondo a luglio. Quest'anno si proverà a fare l'esame al computer/dispositivo tramite moodle. Questo metodo vale per la parte di strutturale (svolta dalla prof.ssa Aldieri), metabolica (svolta dalla prof.ssa Aldieri e Riganti) e d'organo (svolta dalla prof.ssa Riganti) che si svolgerà tramite quiz. Seguirà poi una prova orale. Di solito ai primi due appelli si è sempre tanti e, per far Meisenberg-Simmons. Principi di biochimica medica. Minerva Medicapassare tutti lo stesso giorno, con il pc la correzione dei quiz è automatica e, chi è sufficiente, passa subito all'orale. Se ci fossero comunque troppi iscritti si passerebbe di orale il giorno successivo. Se qualcuno avesse problemi con i dispositivi, può avvisare prima, e la prof.ssa preparerà delle copie cartacee. La parte del professor Turrini consiste in 2/3 domande in modalità scritto cartaceo.
Questa modalità è solo ipotesi, verrà eventualmente confermata più avanti.
SISTEMA NERVOSO CENTRALE
L'argomento verrà svolto in tre lezioni: le prime due sulla fisiologia, la terza verterà su basi biochimiche correlate a patologie come ictus, patologie croniche, Alzheimer ...
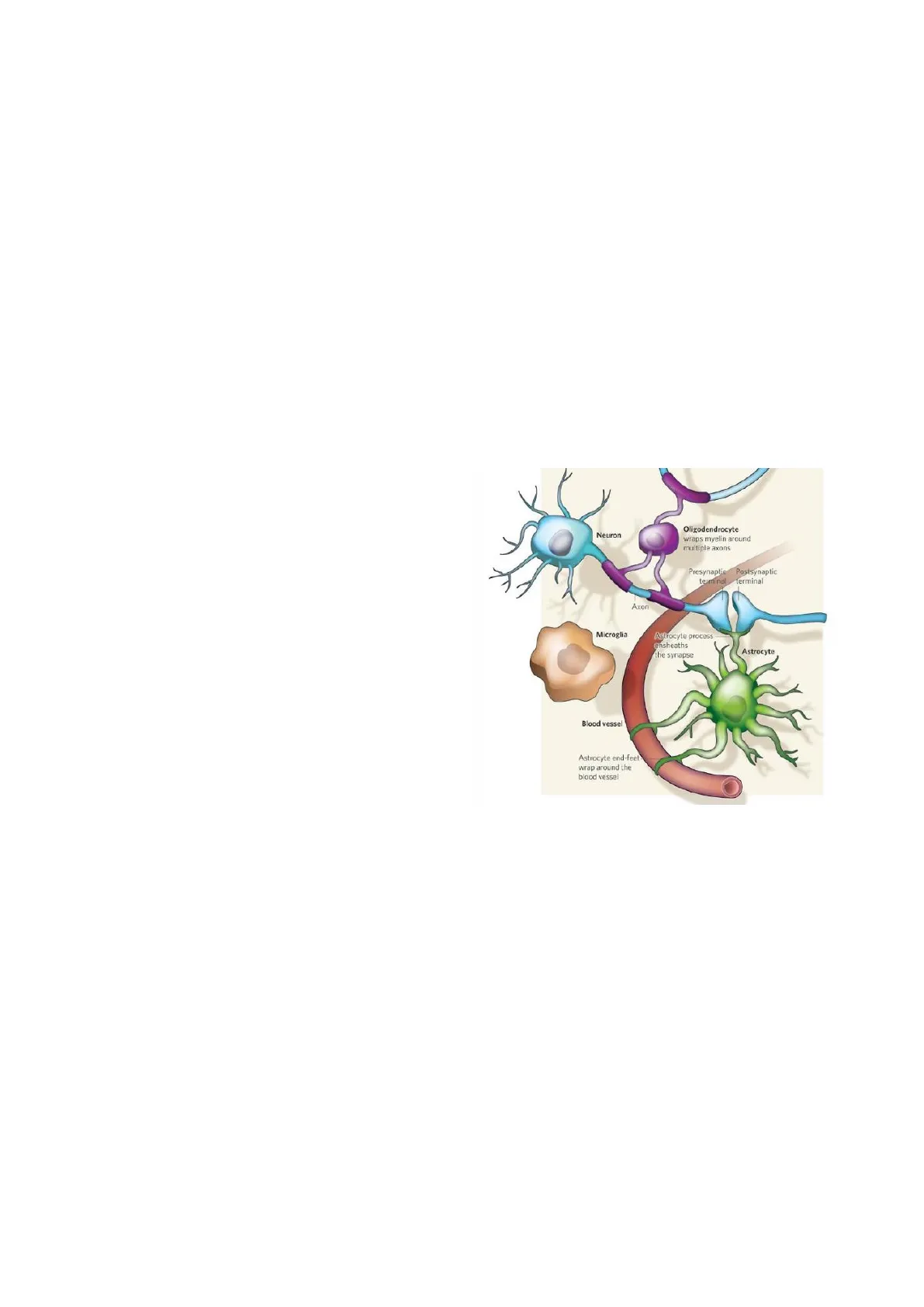
Ci sono 2 grosse tipologie di cellule nel sistema nervoso centrale:
- le cellule neuronali presenti in una quantità di circa 100 bilioni;
- le cellule della glia, con un ordine di grandezza in più, ovvero nell'ordine dei trilioni. Queste, a loro volta, sono divise in:
- astroglia, le cellule si supporto;
- oligodendrociti, sono cellule decretate nella formazione delle guaine mieliniche;
- microglia, cioè i macrofagi del tessuto, e intervengono nelle reazioni dell'infiammazione.
Struttura dei Neuroni
I neuroni si connettono tra di loro e la loro struttura è fatta da un corpo, cioè lo stroma; dei prolungamenti detti neuriti, con cui prendono contatto con le altre cellule; un assone, che è il prolungamento più lungo e che termina con un bottone sinaptico da cui si rilasciano dei neurotrasmettitori.
Consumo Energetico del Cervello
Il cervello è un organo abbastanza piccolo perché rappresenta solo il 2% del peso del corpo, circa 1-1,3 kg, ma ha un consumo energetico altissimo:
- circa il 15% della gittata cardiaca è consumato dal cervello;
- consumo del 20% di O2 (circa 1/5 dell'ossigeno);
- il 25% del glucosio (1/4 del glucosio è richiesto dal cervello, anche in condizioni di riposo, indipendentemente dall'attività): significa che 100-130 g/die vengono consumanti solo dal cervello.
Bisogna considerare che, nonostante la grande dipendenza dal glucoso, le scorte di glicogeno nel cervello sono bassissime e il glucoso rimane l'unica fonte (si vedrà che i neuroni hanno una bassa capacità di ossidare gli acidi grassi). Quindi, il loro funzionamento dipende dall'accumulo di glucosio dalla perfusione.
Meccanismi di Compensazione Cerebrale
Il cervello cerca di attuare dei meccanismi di compensazione: quando sente che c'è una pressione parziale di ossigeno che scende, un'acidificazione e un aumento della pressione parziale di CO2 c'è un rilascio di una serie di mediatori che causano una vasodilatazione dei vasi cerebrali in modo da far arrivare più ossigeno e glucosio.
Al contrario, se tutto funziona bene, si ha una buona pressione parziale di ossigeno, una bassa pressione parziale di CO2 e non c'è acidificazione, allora il cervello rilascia una serie di mediatori che causano una vasocostrizione per risparmiare ossigeno e glucoso perché percepisce che la sua richiesta è già "coperta".
Metabolismo Energetico del Cervello
Di norma, un cervello che è in un buono stato di alimentazione utilizza il glucoso come fonte principale di energia, sia il glucoso che arriva dal sangue (nel periodo post-prandiale, quando immetto del glucoso) sia il glucoso che deriva dal glicogeno degli epatociti. Quest'ultimo viene demolito, immesso nella circolazione; quindi, nel periodo tra un pasto e quello successivo (soprattutto nelle ore notturne) il cervello funziona con il glucoso che deriva dagli epatociti. Quando il digiuno è prolungato, allora si adatta all'uso dei corpi chetonici (l'unica forma di grassi che può sfruttare).
Il metabolismo è sempre aerobio: quindi, il glucoso segue la glicolisi fino al piruvato, che passa poi nei mitocondri, si trasforma in Acetil-CoA, arriva nel ciclo di Krebs e termina con la respirazione cellulare. La stessa cosa si ha con i corpi chetonici che generano dell'Acetil-CoA che passa nel ciclo di Krebs e poi nella respirazione.
Il motivo per cui il cervello utilizza queste vie è perché sono le vie che generano la maggior parte di ATP, rispetto alla glicolisi in anaerobiosi. Questo perché i neuroni sono cellule che hanno necessità di mantenere il potenziale di membrana ben controllato, cioè il gradiente del sodio e del potassio, fisso. Per questo, hanno una grandissima quantità di Na+/K+ ATPasi che pompa il sodio contro gradiente di concentrazione dall'interno della cellula verso l'esterno, e il potassio contro gradiente di concentrazione dall'esterno della cellula all'interno: il tutto a spese di ATP; quindi, c'è un consumo di ATP grande e costante. Il mantenimento del gradiente è indispensabile per il rilascio di neurotrasmettitori, per i meccanismi di trasporto dei neurotrasmettitori, il trasporto degli organelli dal soma al bottone sinaptico ...
Perché gli Acidi Grassi non possono essere Sfruttati?
Perché gli acidi grassi, che sarebbero costanze che produrrebbero molto più ATP, non possono essere sfruttati?
Non è un problema di passaggio, perché si vedrà (a fine lezione) che il cervello è protetto da un endotelio detto barriera ematoencefalica (blood brain barrier = BBB) che regola molto il passaggio delle sostanze, ma possono passare le sostanze lipofile. Quindi, teoricamente i grassi, regolati dall'albumina, potrebbero passare per diffusione. Ma, una volta che arrivano nelle cellule neuronali si trovano davanti delle cellule (i neuroni, appunto) con una bassissima espressione degli enzimi della beta-ossidazione e hanno alta espressione di tutti gli enzimi che servono per la sintesi dei lipidi complessi (fosfolipidi, sfingolipidi ... ). Quindi, quando un acido grasso arriva in una cellula neuronale, viene coniugato e viene inserito nella sintesi di fosfolipidi e di sfingolipidi (per la sintesi delle membrane). Anche le membrane sono importanti perché contribuiscono a mantenere un isolamento elettrico e collaborano a mantenere il potenziale di membrana.
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
