Metabolismo de Lípidos: Catabolismo de Triacilglicéridos y Ácidos Grasos
Diapositivas de Universidad Europea sobre Metabolismo de Lípidos. El Pdf, un material de Biología para Universidad, explica el catabolismo de triacilglicéridos y ácidos grasos, incluyendo la beta-oxidación y el cálculo del rendimiento energético.
Ver más36 páginas


Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Metabolismo de Lípidos
Ue 11
Metabolismo de Carnitina y Ácidos Grasos Esenciales
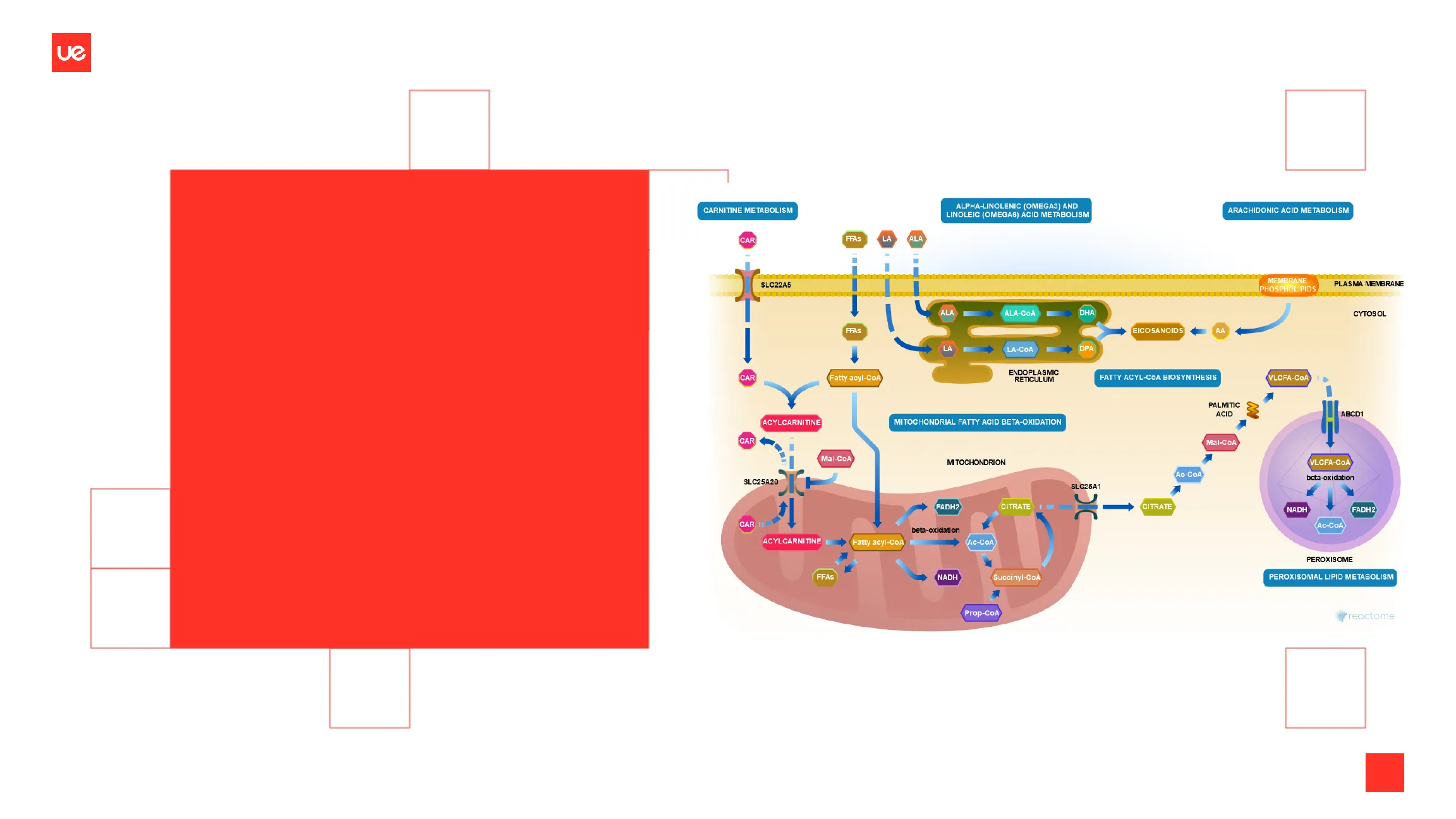
CARNITINE METABOLISM ALPHA-LINOLENIC (OMEGA3) AND LINOLEIC (OMEGA6) ACID METABOLISM ARACHIDONIC ACID METABOLISM CAR FFAS LA ALA SLC22A5 MEMBRANE PHOSPHOLIPIDS PLASMA MEMBRANE ALA ALA-COA DHA CYTOSOL FFAS EICOSANOIDS AA LA LA-COA DPA CAR Fatty acyl-CoA ENDOPLASMIC RETICULUM FATTY ACYL-COA BIOSYNTHESIS VLCFA-COA PALMITIC ACID ABCD ACYLCARNITINE MITOCHONDRIAL FATTY ACID BETA-OXIDATION CAR Mal-CoA VLCFA-COA SLC25A20 SLC25A1 FADH2 CITRATE CITRATE NADH FADH2 CAR 110 beta-oxidation ACYLCARNITINE Fatty acyl-CoA Ac-COA PEROXISOME PEROXISOMAL LIPID METABOLISM FFAS NADH Succinyl-CoA Prop-CoA reactome @ Copyright Universidad Europea. Todos los derechos reservados Mal-CoA MITOCHONDRION Ac-COA beta-oxidation Ac-COAUe
CONTENIDOS
- Introducción.
- Catabolismo de triacilglicéridos.
- Catabolismo de ácidos grasos.
- Anabolismo de triacilglicéridos.
- Metabolismo de los cuerpos cetónicos.
Internal use 2Ue
1. INTRODUCCIÓN
Definición y Enfoque del Metabolismo de Lípidos
El metabolismo de los lípidos es la síntesis y degradación H2C-0-C-CH2-CH2-CH2-CH2-CH3 0 H2C-0-C-CH2-CH2-CH2-CH2-CH3 0 0 H2C-0-C-CH2-CH2-CH2-CH2-CH3 de los lípidos en las células. No es posible estudiar el metabolismo de todos los lípidos ya que muestran diferentes propiedades químicas, luego nos centraremos en el metabolismo de las grasas y los ácidos grasos.
Funciones Fisiológicas de los Ácidos Grasos
Los ácidos grasos tienen cuatro funciones fisiológicas principales: 1) Moléculas de combustible: durante el descanso o el ejercicio moderado, como caminar, los ácidos grasos son nuestra principal fuente de energía. 2) Bloques de construcción: de fosfolípidos y glucolípidos. 3) Modificadores de proteínas: muchas proteínas se modifican por la unión covalente de los ácidos grasos, que dirigen las proteínas a las ubicaciones de las membranas. 4) Mensajeros: los derivados de ácidos grasos sirven como hormonas y mensajeros intracelulares. Internal use 3Ue
1. INTRODUCCIÓN
Regulación del Metabolismo de Ácidos Grasos
El metabolismo de los ácidos grasos está estrictamente controlado, de modo que la síntesis y la degradación responden altamente a las necesidades fisiológicas. La síntesis de ácidos grasos es máxima cuando los carbohidratos y la energía son abundantes y cuando los ácidos grasos son escasos. La enzima acetil CoA carboxilasa regula el metabolismo de los ácidos grasos y responde a los cambios en su entorno inmediato. La acetil CoA carboxilasa está controlada por las hormonas glucagón, epinefrina e insulina, que denotan el estado general de energía del organismo. La actividad de la carboxilasa es estimulada por la insulina e inhibida por el glucagón y la epinefrina. Por lo tanto, la insulina estimula la síntesis de ácidos grasos activando la carboxilasa, mientras que el glucagón y la epinefrina tienen el efecto inverso. Internal use 4Ue
1. INTRODUCCIÓN
Triacilgliceroles como Depósitos de Energía
Los triacilgliceroles son depósitos altamente concentrados de energía metabólica porque son reducidos y anhidros. En mamíferos, el sitio principal de acumulación de triacilglicerol es el citoplasma de las células adiposas (células grasas). La mayoría de los lípidos se ingieren en forma de triacilgliceroles y deben degradarse a ácidos grasos para su absorción a través del epitelio intestinal. Los lípidos presentan un problema especial porque, a diferencia de los carbohidratos y las proteínas, estas moléculas no son solubles en agua. Es por ello que el proceso de absorción requiere varios pasos de interconversión entre triacilgliceroles y ácidos grasos libres. Tras varios pasos de degradación y síntesis para su absorción y transporte, los triacilgliceroles se vuelven a sintetizar dentro de las células adiposas y se almacenan. En el músculo, se pueden oxidar para proporcionar energía. Adipocytes (white adipose cells) Adipose Tissue 5Ue
2. CATABOLISMO DE TRIACILGLICÉRIDOS
Etapas del Procesamiento de Lípidos
Los tejidos obtienen acceso a las reservas de energía lipidica almacenadas en el tejido adiposo a través de tres etapas de procesamiento: 1) Movilización: los lípidos deben ser movilizados. En este proceso, los triacilgliceroles se degradan a ácidos grasos y glicerol, que se liberan del tejido adiposo y se transportan a los tejidos que requieren energía. 2) Activación: en estos tejidos, los ácidos grasos deben activarse y transportarse a las mitocondrias para su degradación. Acetyl COA SCOA palmitoyl Co.A (C16:0) 3) Degradación: los ácidos grasos se descomponen paso a paso en acetil CoA, que luego se procesa en el ciclo del ácido cítrico. `SCEA Acyl CoA FAD+ mysistayt Coà (C11:)) 3-ketoacyl coA thiolase FADH Acyl coA dehydrogenase 3-ketoacyl coA 2-trans-enoyl-coA 0 0 NADH 3-hydroacyl coA dehydrogenase Enoyl-coA hydratase CH O SCO H20 NAD+ (3R)-hydroxyl acyl coA 6Ue
2. CATABOLISMO DE TRIACILGLICÉRIDOS
Conversión de Glicerol en Glucólisis y Gluconeogénesis
H O- H-C- H- C-O- H-C-OH O - O -I H O H-C-OH H O Glicerol Ácido Graso D-Gliceraldehido 3-fosfato Glucólisis ß-oxidación Internal use 7 TAG H H-C-OH O H-C~ -0-Ue
2. CATABOLISMO DE TRIACILGLICÉRIDOS
Metabolismo del Glicerol en el Hígado
El glicerol formado por la lipólisis es absorbido por el hígado y fosforilado. Luego se oxida a fosfato de dihidroxiacetona, que se isomeriza a gliceraldehído 3- fosfato. Esta molécula es un intermediario tanto en la ruta glucolítica como en la gluconeogénica. Por lo tanto, el glicerol puede convertirse en piruvato (glucólisis) o glucosa (gluconeogénesis) en el hígado, que contiene las enzimas apropiadas. Glucosa Glucosa 6-Fosfato Fructosa 6-Fosfato Gliceraldehido 3-Fosfato Dihidroxiacetona Fosfato 1,3-Difosfoglicerato 3-Fosfoglicerato 1 Glicerol Fosfato 1 Glicerol -> 2-Fosfoglicerato Fosfoenolpiruvato 1 Lactato > Piruvato co, 2 Oxaloacetato Aminoácidos glucogenicos Internal use 8Ue
3. CATABOLISMO DE ÁCIDOS GRASOS
Oxidación de Ácidos Grasos en la Matriz Mitocondrial
La degradación de los ácidos grasos es un proceso oxidativo que convierte un ácido graso en un conjunto de unidades de acetilo activadas (acetil CoA) que pueden ser procesadas por el ciclo del ácido cítrico. La oxidación de los ácidos grasos ocurre en la matriz mitocondrial, pero para ingresar a las mitocondrias, los ácidos grasos se activan primero mediante la formación de un enlace tioéster con la coenzima A El proceso se llama ß-oxidación debido a que consiste en la oxidación del carbon beta (C3) a un grupo carbonilo (C=O): Internal use CH3 - (CH2) - CH2 - CH2 - COOH ω β α 9Ue
3. CATABOLISMO DE ÁCIDOS GRASOS
Reacciones de la Beta-Oxidación
Paso Reacción 1 Ácido graso + CoA + ATP -> acil CoA + AMP + PP 2 Carnitina + acil-CoA < > acil carnitina + CoA 3 acil CoA +E-FAD -> trans-42-enoil CoA + E-FADH2 Cíclico 4 trans-42-enoil CoA + H2O <> L-3-hidroxiacil CoA 5 L-3-hidroxiacil CoA + NAD+ <> 3-cetoacil CoA + NADH + H+ 6 3-cetoacil CoA + CoA < > acetil CoA + acil CoA (con dos carbonos menos) Internal useUe
3. CATABOLISMO DE ÁCIDOS GRASOS
Esquema de Reacciones de la Beta-Oxidación
Paso FAD FADH2 3 CoA R -CoA S Acyl-CoA trans-42-Enoyl-CoA OH O + H2O R CoA R CoA S - H2O S trans-42-Enoyl-CoA Enoyl-CoA- Hydratase L-3-Hydroxyacyl-CoA NADH + H+ OH R CoA R CoA S Hydroxyacyl-CoA- Dehydrogenase S L-3-Hydroxyacyl-CoA 3-Ketoacyl-CoA 6 CoA R -COA + -COA S Thiolase S H3C S 3-Ketoacyl-CoA Acyl-CoA Acetyl-CoA Internal use 4 NAD+ W. 5 + CoA-SH R R S Acyl-CoA- DehydrogenaseUe
3. CATABOLISMO DE ÁCIDOS GRASOS
1) Activación del Ácido Graso
O R-CH2-CH,-C-OH + 1) Activación del ácido graso: El equivalente de dos moléculas de ATP se hidroliza para iniciar el proceso: TIOQUINASA ATP Mg++ AMP + PP; + 2 Pi RCOO- + CoA + ATP + H O -> RCO-COA + AMP + 2 P. + 2 H+ 0 R-CH2-CH2-C-S-COA
2) Transporte de Ácidos Grasos Activados por Carnitina
2) La carnitina transporta ácidos grasos activados de cadena larga a la matriz mitocondrial: Acil CoA Los ácidos grasos se activan en la membrana mitocondrial externa, mientras que se oxidan en la matriz mitocondrial. Se necesita un mecanismo de transporte especial para transportar los ácidos grasos de cadena larga activados a través de la membrana mitocondrial interna. Estos ácidos grasos deben conjugarse con carnitina para formar acilcarnitina. CoA-SH SA Pirofosfatasa Internal useUe
3. CATABOLISMO DE ÁCIDOS GRASOS
Mecanismo de Entrada de Acilcarnitina en la Matriz Mitocondrial
La entrada de acilcarnitina en la matriz mitocondrial está mediada por una translocasa (una proteína de membrana). La carnitina regresa al lado citoplasmático de la membrana mitocondrial interna a cambio de acilcarnitina. Acil CoA CoA Citoplasma Acilcarnitina Carnitina Translocasa Carnitina Acilcarnitina Acil CoA CoA Matriz mitocondrial Internal useUe
3. CATABOLISMO DE ÁCIDOS GRASOS
Transporte de Ácidos Grasos a la Matriz Mitocondrial
Cytosol Outer Mitochondrial Membrane Inner Mitochondrial Membrane Mitochondrial Matrix R Fatty acid OH Acyl CoA CoA-SH synthetase ATP Intermembrane space Kreb's cycle - Acetyl COA ß-oxidation AMP + PP; Carnitine acyltransferase II Fatty Acyl-CoA O Transporter/ Carrier R R- S-CoA Carnitine S-CoA R O Carnitine R CoA-SH CoA-SH Fatty Acyl- Carnitine Fatty Acyl- Carnitine Carnitine Carnitine acyltransferase I Pharma Change .info 1 Fatty Acyl-CoA O Carnitine OUe
3. CATABOLISMO DE ÁCIDOS GRASOS
3) Beta-Oxidación y Rendimiento Energético
3) ß-oxidación: Un acil CoA saturado se degrada por una secuencia recurrente de cuatro reacciones. La cadena de ácido graso se acorta por dos átomos de carbono como resultado de estas reacciones y se generan FADH2, NADH y acetil CoA en cada ciclo. El número de ciclos (N) de ß-oxidación por ácido graso de n átomos de carbono es: N ==- 1 2 n Si n = 16 (ácido palmítico) -> N = (16/2) - 1 = 7 ciclos. El número Acetil-CoA en la ß-oxidación por ácido graso de n átomos de carbono es: Acetil-CoA=n/2 Si n = 16 (ácido palmítico) -> nº Acetil-CoA = (16/2) = 8 Acetil-CoA. Internal useUe
3. CATABOLISMO DE ÁCIDOS GRASOS
Cálculo de Ciclos y Acetil-CoA para Ácido Mirístico
Si n = 14 (ácido mirístico) -> N = (14/2) - 1 = 6 ciclos -> N = (14/2) = 7 Acetil-CoA C14 Acetil CoA Acetil CoA C12 Acetil CoA C10 Ciclo de Krebs > Acetil CoA C3 C6 Acetil CoA Acetil CoA C4 C2 Acetil CoA NADH + FADH2 Cadena de transporte de electrones Internal use 94 ATPUe
3. CATABOLISMO DE ÁCIDOS GRASOS
Rendimiento Energético de la Oxidación de Ácidos Grasos
Ahora podemos calcular el rendimiento energético derivado de la oxidación de un ácido graso. En cada ciclo de reacción, un acil CoA se acorta con dos átomos de carbono, y se forma una molecula de FADH2, NADH y acetil CoA. C-acil CoA + FAD + NAD+ + CoA + H2O -> Cn-2-acil CoA + FADH2 + NADH + H+ + Acetil CoA + H+ Por ejemplo, la degradación de palmitoil CoA (C16-acil CoA) requiere siete ciclos de reacción. En el séptimo ciclo, el C4-cetoacil CoA se transforma en dos moléculas de acetil CoA. Por lo tanto, la estequiometría de la oxidación de palmitoil CoA es: Palmitoil CoA + 7 FAD + 7 NAD+ + 8 CoA + 7 H2O -> 8 acetil CoA + 7 FADH2 + 7 NADH + 7 H+ La oxidación completa del palmitato (C16) produce 106 moléculas de ATP (teóricamente). Internal use
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
