Dominio Archaea: características generales, hipertermófilos, halófilos y metanógenos
Documento de Universidad sobre Dominio Archaea: Características Generales, Hipertermófilos, Halófilos y Metanógenos. El Pdf, de Biología, explora la composición de la membrana celular, la pared celular y los metabolismos específicos, incluyendo géneros como Sulfolobus y Thermoproteus.
Ver más19 páginas

Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Características Generales de las Archaea
Tema 2 .- Dominio Archaea. Características generales. Hipertermófilos, Halófilos y Metanógenos. A finales de los años 70 se propuso una teoría revolucionaria: Los procariotas no forman un grupo monofilético, sino que existen dos dominios diferenciados que denominaron bacterias y Archaeas. Las Archaeas constituyen un grupo bien diferenciado teniendo la siguientes características:
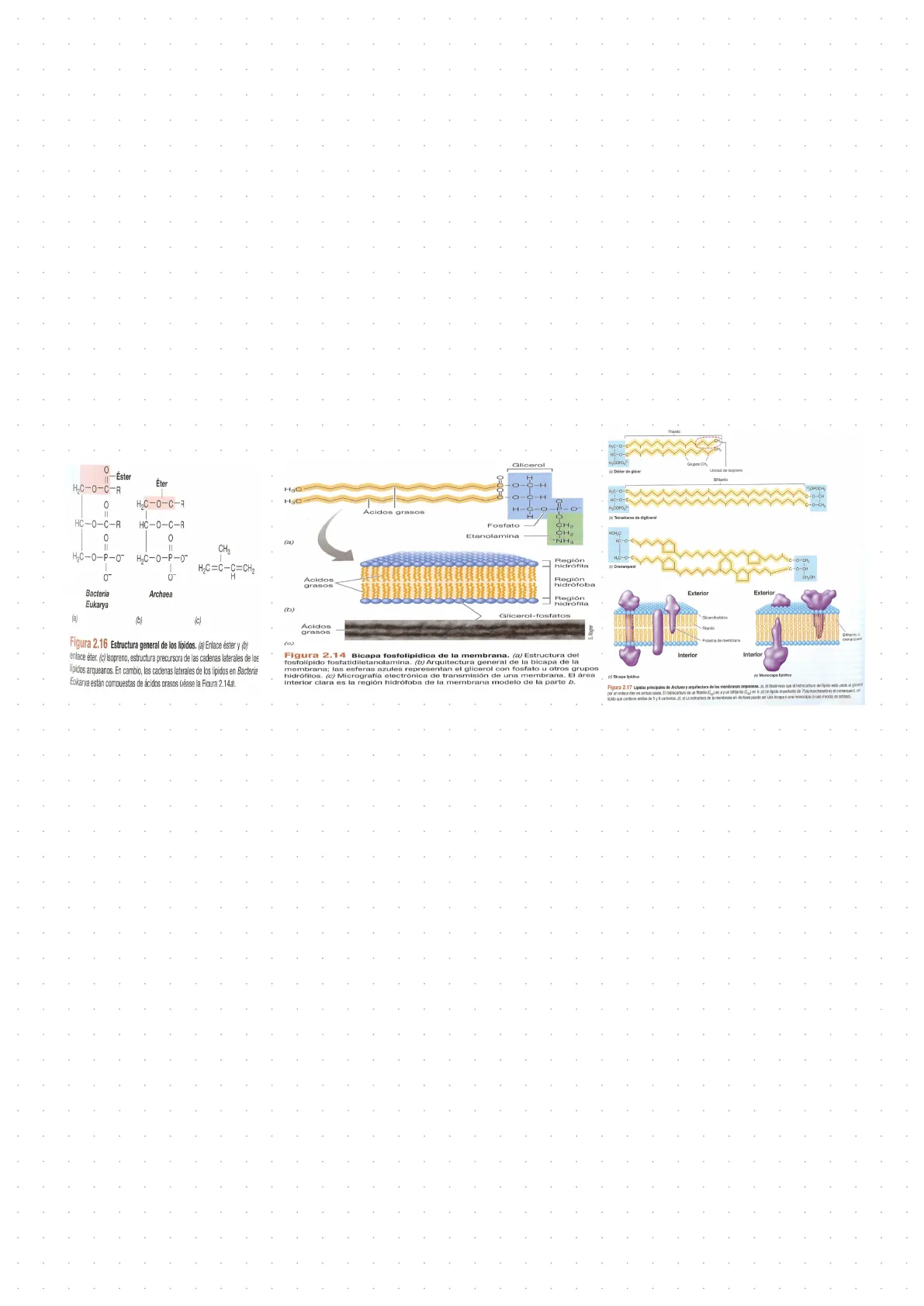
- Los lípidos de membrana tienen enlaces eter (mientras que los miembros de bacteria y Eukarya tienen enlaces ester). Las arqueas pueden formar varios tipos de lípidos: dieteres de glicerol, que se forman con el fitanilo normalmente que es derivado del isopreno, que formaría una bicapa lipídica; pero también puede formar tetraeteres de diglicerol, que es este caso el hidrocarburo tiene un enlace éter por cada lado, y en este caso el hidrocarburo es el bifitanilo. Estos tetraeteres de diglicerol forman monocapas lipídicas. Otro tipo de lípidos que podemos encontrar en las Arqueas se denomina Crenarqueol, y se encuentran exclusivamente en el filo Thaumarchaeota. Fitanilo 0 -Éster Éter H2C-0-C-R 0 H2C-0-C-R HC-0-C-R HC-0-C-R 0 0 II CH3 H2C-0-P-0" H2C-0-P-0" 1 H2C=C-C=CH2 H Bacteria Eukarya Archaea (a) (b) (c) Figura 2.16 Estructura general de los lípidos. (a) Enlace éster y (b) enlace éter. (c) Isopreno, estructura precursora de las cadenas laterales de los lípidos arqueanos. En cambio, las cadenas laterales de los lípidos en Bacteria Eukarya están compuestas de ácidos grasos (véase la Fioura 2.14a). H.C-O HC-0- H2COPO, Grupos CH3 (o) Dieter de glicer Unidad de isoprono Biftanilo ",OPOCH, LC-0-C C-O-CH HC-0-C HyCOPO- b/ Tetraéteres de diglicerol HOH,C HC-O-C HC-Q-C C-O-CH2 c) Crenarqueol C-O-CH CH-CH Exterior Exterior Gliceratostatos Fitanilo Bifibanilo @ Proteina de membrana crenarqueol Interior Interior : (c) Bicapa lipidica je) Monocapa lipidion Figura 2.17 Lipides principales de Archaea y arquitectura de las membranas arqueanas. (a, b) Obsérvese que el hidrocarburo del lipido está unido al glicerol por un enlace éter en ambos casos. El hidrocarburo es un fitanilo (C ) en ay un bifitanilo (Ca) en b. (c) Un lipido importante de Thaumarchanota es el crenarqueol, un lipido que contiene anillos de 5 y 6 carbonos. (d, e) La estructura de la membrana en Archaea puede ser una bicapa o una monocapa (o una mezcla de ambas).
- La pared celular no contiene peptidoglicano, sino que tienen una mayor diversidad de paredes celulares. La pared celular más frecuente es la capa S que es una estructura externa a la membrana plasmática y paracristalina formada por proteínas o glicoproteínas. Esta estructura contendría la membrana plasmática y por encima la capa S. Es decir que su estructura sería; capa S + mp. Luego hay otros que pueden ser variaciones de la capa S o capas de composiciones diferentes, por ejemplo:
- Hay Archaeas que contienen metanocondroitina en su pared celular (aparece en algunos metanogenos) es un heteropolímero de N-acetil-galactosamina más N- glucurónico. Estas Archaeas tienen una estructura de Metanocondroitina por el exterior, luego esta la capa S y por último la membrana plasmática.
- Hay Archaeas que tienen un polímero en la pared celular similar a la mureína que se denomina pseudomureína o pseudopeptidoglucano la característica de este polímero es que no tiene N-Acetil murámico, en su lugar tiene N-acetil- talosaminurónico + N-acetil-glucosamina, donde sus uniones son diferentes (beta- 1,3). Además, no tiene D- aminoácidos en las uniones intrapeptidicas, pero tiene L- aminoácidos. Su estructura es : Capa S + Pseudomureína + mp.
- Hay Archaeas que tienen una pared celular constituidas por una capa gruesa de polisacárido complejo que puede tener una composición variable, por lo que tendría un polisacárido por encima de la pared plasmática.
- Por último, hay Archaeas sin pared celular como por ejemplo los Termoplasmas. G. Wagner (c) Figura 2.14 Bicapa fosfolipidica de la membrana. (a) Estructura del fosfolipido fosfatidiletanolamina. (b) Arquitectura general de la bicapa de la membrana; las esferas azules representan el glicerol con fosfato u otros grupos hidrófilos. (c) Micrografía electrónica de transmisión de una membrana. El área interior clara es la región hidrófoba de la membrana modelo de la parte b. Glicerol 0=00=0 H C-H H3C HJC Ácidos grasos H-C .O 0=1- 0 Fosfato Etanolamina CH. CH2 +NH3 (a) Región hidrófila Región hidrofoba Región hidrófila (b) Glicerol-fosfatos Ácidos grasos 0 0 -C-H -0-CH2 -0 Ácidos grasos· Su RNA polimerasa tiene una estructura más parecida a eucariotas, por la cantidad de subunidades, que a la de bacterias.
Filogenia y Diversidad Metabólica de Archaea
Actualmente el dominio Arquea se divide en 5 filos: Euryarchaeota, Nanoarchaeota, Korarchaeota, Crenarchaeota y Thaumarchaeota. Se establecen en base al ADN ribosomal 16S. Estos 5 filos se pueden organizar en 4 grandes superfilos: Euryarchaeota, Grupo TACK, Asgard o Asgardarchaeota y Grupo DPANN. Presentan una gran diversidad fenotípica, hay especies con metabolismo quimiorganotrofo donde oxidan materia orgánica para obtener energía, y que está muy extendida; pudiendo ser aerobia y anaerobia. En el caso de la anaerobia puede estar asociado a la respiración anaerobia o incluso a la fermentación. También puede haber metabolismo quimiolitotrofo que oxidan compuestos inorgánicos para obtener la energía. Un donador frecuentes de electrones en este ultimo tipo es el Hidrógeno. Dentro de las arqueas hay características metabólicas únicas como la metanogénesis: que es la producción de metano como la consecuencia del metabolismo energético, proceso que solo llevan algunas arqueas, y es muy importante a escala global en el efecto invernadero. Muchas son extremofilas, que viven donde se consideran los limites de la vida. Muchas son hipertermófilas (la mayoría de las arqueas, temperatura óptima de crecimiento a los 80 grados o más, donde su velocidad de crecimiento es la máxima posible), otras halófilas extremas (crecen con sal), incluso acidofilos extremos (crecen con un pH muy bajo, mucha acidez). Pero no todas son extremófilas.
Árbol Filogenético del Dominio Archaea
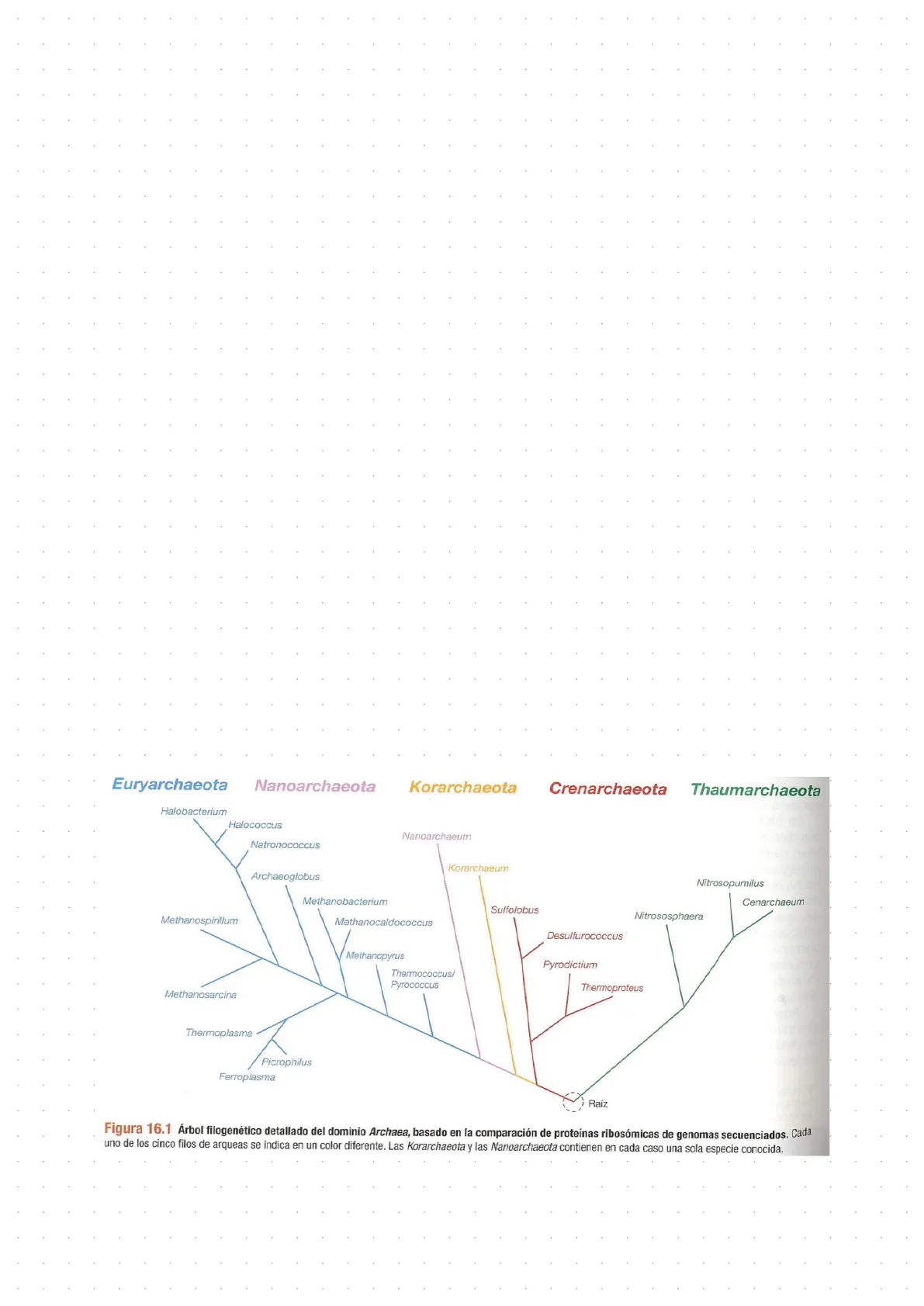
Árbol filogenético del dominio Archaea Methanobacteriota Euryarchaeota "Candidatus" Nanoarchaeota "Candidatus" Korarchaeota Thermoproteota Crenarchaeota Nitrososphaerota Thaumarchaeota Halobacterium Halococcus Nanoarchaeum Natronococcus Archaeoglobus Nitrosopumilus Methanobacterium Cenarchaeum Sulfolobus Nitrososphaera Methanospirillum Methanocaldococcus Desulfurococcus Methanopyrus Pyrodictium Thermococcus/ Pyrococcus Thermoproteus Methanosarcina Thermoplasma Picrophilus Ferroplasma Raíz Figura 16.1 Árbol filogenético detallado del dominio Archaea, basado en la comparación de proteínas ribosómicas de genomas secuenciados. Cada uno de los cinco filos de arqueas se indica en un color diferente. Las Korarchaeota y las Nanoarchaeota contienen en cada caso una sola especie conocida. KorarchaeumDominio Archaea
Superfilos de Archaea
Superfilo Euryarchaeota Grupo TACK Thaumarchaeota Korarchaeota Crenarchaetoa Candidatus Aigarchaeota Candidatus Bathyarchaeota Candidatus Geoarchaeota Asgard (Asgardarchaeota) Candidatus Lokiarchaeota Candidatus Odiarchaeota Candidatus Thorarchaeota Candidatus Heimdallarchaeota Grupo DPANN Nanoarchaeota Candidatus Diapherotrites Candidatus Parvarchaeota Candidatus Micrarchaeota Candidatus Woesearchaeota Candidatus Pacearchaeota Candidatus Aenigmarchaeota Candidatus Nanohaloarchaeota Dominio Archaea
Phylum Thermoproteota Crenarchaeota
Phylum Thermoproteota Crenarchaeota ¡OJO! En las clasificaciones posteriores nos referimos a Filo Thermoproteota. Dominio Archaea Phylum Thermoproteota Crenarchaeota Hay muchas Archaeas cultivables (La mayoría de Archaeas cultivables se encuentran en Crenarchaeota y Euryarchaeota). Dentro de las Archaeas cultivables las del phylo Crenarchaeota son las más hipertermófilas, de hecho, hay muchas especies con una temperatura óptima de crecimiento mayor a 100oC (estas condiciones se consiguen en las fuentes hidrotermales porque el agua se mantiene líquida a 100oC), por ello muchos de los hipertermófilos son anaerobios porque el oxígeno se disuelve mal en el agua caliente. Muchos microorganismos que se encuentran a temperaturas superiores a 100oC suelen ser quimiolitoautótrofos porque en estos ambientes la materia orgánica es escasa y porque estas especies suelen ser productores primarios. La mayoría de las Archaeas hipertermófilas se aíslan en suelos o aguas calentados por energía geotérmica y suelen ser ambientes ricos en azufre (A estos ambientes se les denomina solfataras), por tantos los microorganismos de estos ambientes suelen metabolizar el azufre.
- La mayoría de Archaeas cultivables se encuentran en Crenarchaeota y Euryarchaeota.
- Son los más hipertermófilos.
Phylum Crenarchaeota
Dominio Archaea Phylum Crenarchaeota Clase Thermoprotei Muchas están próximas al antecesor universal porque son hipertermófilos (la mayoría de estos están cerca de la raíz) y muchos son termófilos o hipertermófilos. Se cree que las primeras formas de vida eran hipertermófilas. Dominio Archaea Phylum Crenarchaeota Clase Thermoprotei
Orden Sulfolobales
Orden SulfolobalesTodos los miembros del orden Sulfolobales se aíslan de hábitats volcánicos terrestres. Tiene dos géneros: Sulfolobus y Acidianus. Dominio Archaea Phylum Crenarchaeota Clase Thermoprotei
Género Sulfolobus
Orden Sulfolobales Género Sulfolobus Fue el primer microorganismo termófilo que se aisló, en concreto fue Sulfolobus acidocaldarius. Son microorganismos irregularmente esféricos y lobulado. Se aíslan normalmente de manantiales hidrotermales ácidos ricos en azufre donde pueden crecer con temperaturas de hasta 90oC (con temperatura óptima de 70 a 80oC), pueden crecer a un pH entre 1 y 4 pero su pH óptimo está entre 2 y 3, por lo tanto, se dice que son termoacidófilos. Contienen una pared celular compleja formada por proteínas e hidratos de carbono. Son quimiolitoautótrofos, obtienen el carbono por fijación del CO2 a través del ciclo del 3-hidroxipropionato/4- hidroxibutirato. Cuando crecen como quimiolitoautótrofos utilizan como donador de electrones normalmente el azufre (S0) o el sulfhídrico (SH2) que oxidan hasta sulfato. Pueden crecer también como quimiorganotrofo donde oxidan azucares (aminoácidos o digeridos de proteínas). Pueden utilizarse en biominería (recuperan metales de menas de baja ley). Las especies de sulfolobus son relativamente sencillas a la hora de cultivarlas en el laboratorio, se utilizan como modelo de adaptación de los microorganismos a estos ambientes tan extremos, se ha visto que la membrana de la célula de sulfolobus tiene tetraéteres de diglicerol que puede suponer hasta el 98% de los lípidos de su membrana; estos lípidos son altamente impermeables a los protones de manera que a sulfolobus le permiten mantener un pH interno próximo a 6,5.
Biominería con Sulfolobus
BIOMINERÍA Puede oxidar el ion ferroso (Fe2+) hasta ion férrico (Fe3+) y esta propiedad se puede utilizar en biominería. La biominería es una técnica donde se utilizan microorganismos para recuperar metales anclados en menas de baja ley, normalmente estos metales suelen estar unidos a azufre. Hay varias formas en las que se puede hacer, pero sólo es necesario saber que es importante en biominería la obtención del ion férrico (Fe3+) (Que obtiene sulfolobus oxidando ion ferroso (Fe2+)) porque es un potente oxidante de los sulfuros (pirita), al oxidar el azufre de los sulfuros presentes en la pirita forma ácido sulfúrico y libera el ion ferroso (altamente inestable en presencia de oxígeno salvo que esté en medio ácido). Por lo tanto, si tenemos un metal unido a la pirita lo podemos hacer reaccionar con el ion férrico que oxida el azufre liberando el metal.
Resumen de Sulfolobus
RESUMEN
- Irregularmente esféricos y lobulados.
- Contiene tetraéteres de glicerol.
- Viven en manantiales hidrotermales ricos en azufre.
- Son termoacidófilos.
- Pared celular compleja (proteínas e hidratos de carbono).
- Quimiolitoautótrofos y quimiorganotrofos.
- Ciclo del 3-hidroxipropionato/4-hidroxibutirato.
- Donador de electrones: el azufre (SO) o el sulfhídrico (SH2). Oxidan hasta sulfato.
- Puede oxidar el ion ferroso (Fe2+) a ion férrico (Fe3+).
- Especie: Sulfolobus acidocaldarius.
- Biominería.
Género Acidianus
Dominio Archaea Phylum Crenarchaeota Clase Thermoprotei Orden Sulfolobales Género Acidianus
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
