Glicolipidi e proteine di membrana: struttura e funzione all'università
Slide dall'Università su Glicolipidi e Proteine di Membrana. Il Pdf esplora la struttura e la funzione di questi componenti cellulari, descrivendo i diversi tipi di lipidi come fosfatidilcolina e sfingomieline, e le proteine integrali e periferiche di membrana, utile per lo studio della Biologia a livello universitario.
Mostra di più65 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Catene Aciliche e Glicolipidi
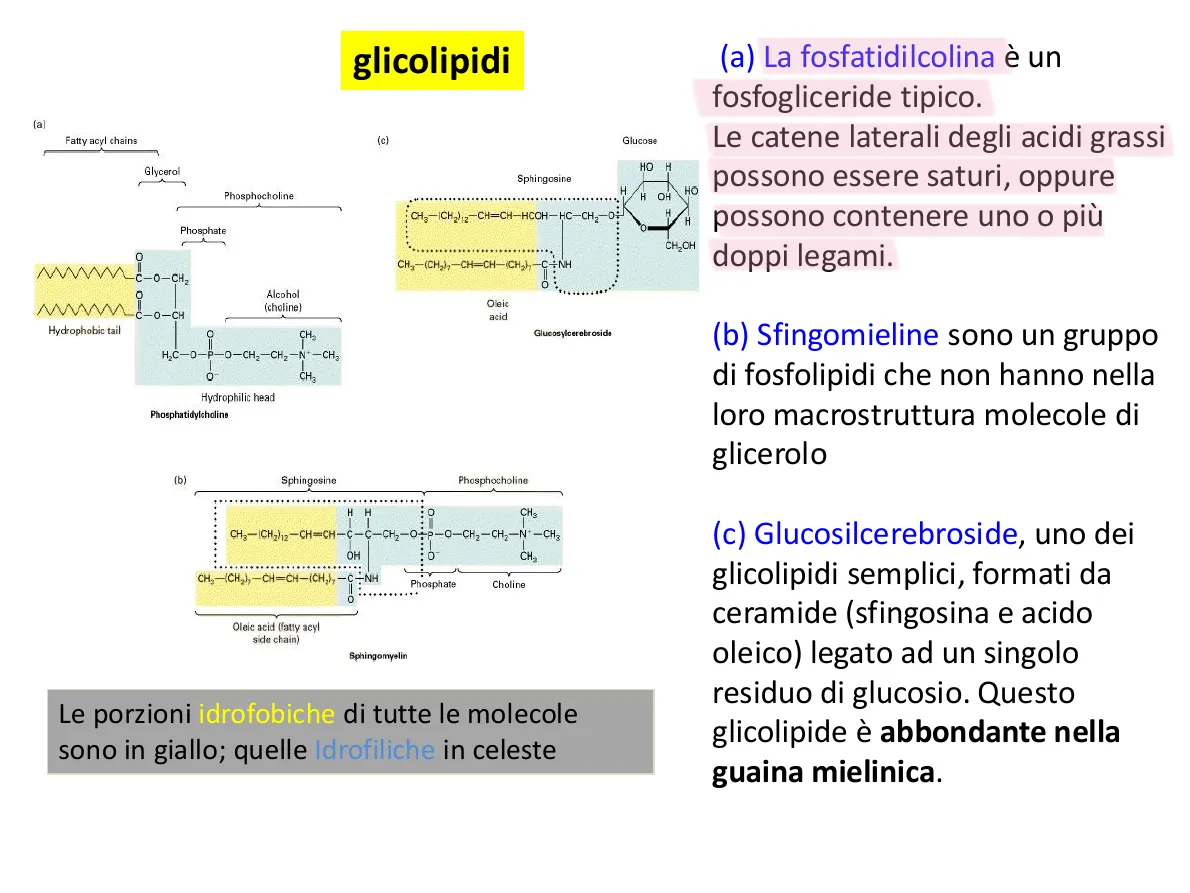
(a)
Fatty acyl chains
(c)
Glucose
HO
H
Sphingosine
H
HO
H
OH
CH2-(CH2)12-CH=CH-HCOH-HC-CH2-0:
H
-I
H
CH,OH
CH2-(CH2),-CH=CH-(CH),-C+NH
ö
Oleic
acid
Glucosylcerebroside
0
CH3
Hydrophilic head
Phosphatidylcholine
(b)
Sphingosine
Phosphocholine
H H
0
CH3
CH3-(CH2)12-CH=CH-C-C-CH2-0:P-O-CH2-CH2-N+-CH3
-
OH
0
CH2
CH2-(CH2),-CH=CH-(CH)),-C+-NH
Phosphate
Choline
0
Oleic acid (fatty acyl
side chain)
Sphingomyelin
Le porzioni idrofobiche di tutte le molecole
sono in giallo; quelle Idrofiliche in celeste
(a) La fosfatidilcolina è un
fosfogliceride tipico.
Glycerol
Phosphocholine
Phosphate
0
-C-0-CH2
O=0
Alcohol
(choline)
C-O-CH
Hydrophobic tail
0
CH2
H2C-0-P-O-CH2-CH2-N+-CH3
Le catene laterali degli acidi grassi
possono essere saturi, oppure
possono contenere uno o più
doppi legami.
(b) Sfingomieline sono un gruppo
di fosfolipidi che non hanno nella
loro macrostruttura molecole di
glicerolo
(c) Glucosilcerebroside, uno dei
glicolipidi semplici, formati da
ceramide (sfingosina e acido
oleico) legato ad un singolo
residuo di glucosio. Questo
glicolipide è abbondante nella
guaina mielinica.
Composizione Lipidica delle Membrane
glicolipidiLa composizione lipidica dei monostrati interno ed esterno della membrana è
diversa.
Questo riflette le diverse funzioni delle due facce di una membrana cellulare.
- Diverse miscele di lipidi si trovano nelle membrane delle cellule di tipo diverso,
così come nelle varie membrane di una singola cellula eucariotica. - Alcuni enzimi di membrana per la loro funzione richiedono i gruppi di testa dei
lipidi. - I gruppi di testa di alcuni lipidi formano siti di attracco per proteine citosoliche
specifiche. - Alcuni segnali extracellulari che agiscono attraverso recettori di membrana
attivano fosfolipasi che degradano molecole selezionate di fosfolipidi nella
membrana plasmatica, generando frammenti che agiscono come molecole di
segnalazione intracellulare.
Struttura delle Membrane Cellulari
Tutte le membrane cellulari sono strutture
chiuse, che circondano la cellula stessa o i
singoli compartimenti.
Le membrane cellulari hanno quindi una
faccia interna (lato orientato verso il
citoplasma detta citosolica) e una faccia
esterna (lato orientato verso l'ambiente).
Poiché la maggior parte degli organelli sono
circondati da una singola membrana a
doppio strato, è anche utile per parlare della
faccia citosolica e exoplasmatica faccia della
membrana, essendo la parte del citoplasma
di fuori degli organelli
Plasma membrane
Golgi
O
X
Nucleus
ER
Mitochondrion
Lysosome
Endosome
Multivesicular body
Membrane bilayer
Exterior
Cytosol
Polar head
groups
Hydrophobic
tails
Polar head
groups
Spazio Chiuso e Doppi Strati Simmetrici
Ogni membrana cellulare forma uno spazio chiuso (lume) che ha una
faccia citoplasmastica e una faccia exoplasmatica
I fosfolipidi nelle cellule formano
spontaneamente doppi strati simmetrici
Le catene laterali di idrocarburi formano un
core idrofobico che è 3 - 4 nm di spessore
Nelle biomembrane i fosfolipidi differiscono
nella carica dei gruppi di testa polari
Esempio:
-fosfatidilcolina e fosfatidiletanolammina non
hanno carica elettrica netta;
-fosfatidilglicerolo e fosfatidilserina hanno una
carica netta negativa.
Polar head groups
Double bond
Hydrophobic interior
Leaflet
1 nm
Polar head groups
Hydrogen -
Carbon
Oxygen
Nitrogen
Phosphorus
I gruppi di testa polari in tutti i fosfolipidi si
dispongono all'esterno delle code idrofobiche
Differenze nella Composizione dei Foglietti Lipidici
La composizione dei fosfolipidi nel doppio strato lipidico differisce
nei due foglietti
La maggior parte dei tipi di fosfolipidi, così come il colesterolo, sono
generalmente presenti in entrambi i foglietti della membrana, anche
se sono spesso più abbondanti in uno o l'altro.
Ad esempio, in membrane plasmatiche di eritrociti umani quasi tutta la
sfingomielina e fosfatidilcolina (lipidi con testa carica positivamente) si
trovano nel foglietto esterno .
Al contrario, lipidi con gruppi di testa neutri o negativi polari (ad
esempio, fosfatidiletanolamina, fosfatidilserina e fosfatidilinositolo)
sono localizzati nel foglietto citosolico.
Movimento dei Lipidi nelle Membrane
La maggior parte dei lipidi si muovono lateralmente nelle membrane
Nel doppio strato di fosfolipidi delle membrane naturali, il moto
termico permette ai fosfolipidi e glicolipidi di ruotare liberamente
attorno ai loro assi e di diffondere lateralmente all'interno del
foglietto della membrana.
Poiché tali movimenti sono laterali o rotazionali, le catene
idrocarburiche degli acidi grassi rimangono all'interno della regione
idrofobica della membrana.
In entrambe le membrane naturali e artificiali, una tipica molecola lipidica
scambia posizione con i suoi vicini circa 107 volte al secondo e diffonde
diversi micrometri per secondo a 37 ° C.
A questo ritmo, un lipide potrebbe diffondere nella lunghezza di una tipica
cellula batterica (~1 micron) in soltanto 1 secondo e la lunghezza di una
cellula animale in circa 20 secondi.
Movimento Flip-Flop dei Lipidi
Oltre al movimento laterale è anche possibile, entro un certo limite, il
passaggio da uno strato all'altro delle singole molecole lipidiche per
inversione testa-coda.
Questo movimento, è detto "flip-flop
Da notare che in doppi strati di fosfolipidi puri, fosfolipidi non migrano,
o flip-flop, da un foglietto della membrana all'altro.
In alcune membrane naturali, tuttavia, lo fanno raramente, grazie a
alcune proteine di membrana chiamate flippasi
Energeticamente, tali movimenti sono estremamente sfavorevoli,
perché la testa polare di un fosfolipide deve essere trasportata
attraverso l'interno della membrana idrofoba.
Le proteine non sono coinvolte nel meccanismo flip-flop attraverso
una membrana.
Morfologia Trilaminare e Tecnica di Congelamento-Frattura
Morfologia trilaminare
La microscopia elettronica di sezioni
sottili della membrana colorate con
tetrossido di osmio, che si lega
fortemente ai gruppi di testa polari di
fosfolipidi, ha fornito la prova più
diretta per l'universalità della struttura
a doppio strato.
Una sezione trasversale di tutte le
singole membrane colorate con
tetrossido di osmio si presenta come un
binario ferroviario: due linee scure
sottili con uno spazio uniforme della
luce di circa 2 nm (le code idrofobiche)
tra loro.
Separazione dei Foglietti Lipidici
(a) Cell or tissue is
frozen in nitrogen
(b) Fracture ruptures
the cell
(c) Integral proteins remain
embedded in fractured leaflets
E (exoplasmic) face
P (protoplasmic) face
La tecnica del congelamento- frattura
consente di separare i due foglietti di
fosfolipidi che formano ogni membrana
cellulare
(a) Una preparazione di cellule o tessuti viene
rapidamente congelato in azoto liquido a -
196 ° C, che immobilizza all'istante
componenti cellulari.
(b) Il blocco delle cellule congelate è
fratturato con un colpo secco da un
coltello freddo. Il piano di frattura è
irregolare.
(c) proteine di membrana e le particelle
rimangono legati ad un foglietto o all'altro,
come illustrato nella figura.
Proteine di Membrana
proteine di membrana
Le due categorie di base
di proteine di membrana
sono:
1-proteine integrali, in
tutto o in parte di che
penetrano o estendono il
doppio strato di
fosfolipidi
2-proteine periferiche,
che non interagiscono
con il nucleo idrofobico
del doppio strato
Peripheral protein
Exterior
Oligosaccharide
Glycoprotein
Glycolipid
800
Integral protein
900
800
800
40
000
Leaflets
Hydrophobic
core
Fatty acyl
tails
Cytosol
Integral protein
Hydrophilic polar
head
Peripheral proteins
Phospholipid
bilayer
Phospholipid
Proteine Integrali di Membrana
Proteine integrali di membrana, dette anche proteine intrinseche, hanno
uno o più segmenti che sono incorporati nel doppio strato fosfolipidico.
La maggior parte delle proteine integrali contengono residui con catene
laterali idrofobiche che interagiscono con i gruppi acilici degli acidi grassi dei
fosfolipidi di membrana, ancorando così la proteina alla membrana.
La maggior parte delle proteine integrali coprono l'intero doppio strato
fosfolipidico.
Queste proteine transmembrana
contengono uno o più domini di
membrana, lunghi da quattro a
diverse centinaia di residui, che
si estendono nel mezzo acquoso
su ciascun lato del doppio strato.
doppio
strato
lipidico
W
3
1
2
4
Ancoraggio delle Proteine Integrali
Alcune proteine integrali sono ancorate ad uno dei foglietti
di membrana di acidi grassi legati covalentemente.
In queste proteine, l'acido grasso legato viene incorporato
nella membrana, ma la catena polipeptidica non entra nel
doppio strato fosfolipidico.
NH2
doppio
strato
lipidico
3
1
2
7
4
Domini Alfa-Elica Transmembrana
Proteine integrali che contengono domini a-elica membrane-spanning
sono incorporati nelle membrane e stabilizzate da interazioni idrofobiche
con l'interno del doppio strato lipidico e da interazioni ioniche con i
gruppi di testa polari dei fosfolipidi.
Es. Una famiglia di
proteine integrali è definita
dalla presenza di sette a-
eliche che attraversano la
membrana.
C-terminus
Cytosol
B
C
A
E
Phospholipid
bilayer
G
F
Retinal
pigment
D
Exterior
N
doppio strato
lipidico
SPAZIO
EXTRA-
CELLULARE
9
PERIPLASMA
(A)
OmpA
a 8 filamenti
(B)
OMPLA
a 12 filamenti
(C)
porina
a 16 filamenti
2 nm
(D)
FepA
a 22 filamenti
1
N-terminus
Proteine Periferiche di Membrana
Proteine di membrana periferiche, o proteine estrinseche, non
interagiscono con il nucleo idrofobico del doppio strato fosfolipidico.
Di solito sono legate alla membrana indirettamente dalle
interazioni con le proteine integrali di membrana o
direttamente da interazioni con i lipidi gruppi di testa polari.
(A)
(B)
(C)
NH-
HO
=
-C-C-O-CH3
CH2
P
P
H-N
5
c=0
c=0
CITOSOL
doppio
strato
lipidico
CITOSOL
(D) ancora miristilica
(E) ancora palmitica
(F) ancora farnesilica
O
1
C-O ...
1
O
o=
COOH
5
3
legame ammidico
fra il gruppo
amminico terminale
e l'acido miristico
legame tioestere
fra cisteina
e gruppo palmitico
S
legame tioetere
fra cisteina
e gruppo prenilico
CH2
CH2 ·····
Ancoraggio di Proteine Integrali di Membrana Plasmatica
Esempi di ancoraggio delle proteine integrali di
membrana plasmatica da parte di gruppi di
idrocarburi immersi nella membrana.
La proteinaThy-1 e altri diversi enzimi
idrolitici sono ancorati al
glicosilfosfatidilinositolo.
Questo ancoraggio si trova solo sulla
faccia exoplasmatica.
Proteine citosoliche coinvolte nella
segnalazione come Ras sono ancorate
alla faccia citosolica della membrana
attraverso gruppi farnesil e palmitoil.
Altre proteine citosoliche sono
associate con la membrana attraverso
miristato e acidi grassi simili, collegati a
un residuo di glicina nella estremità N-
terminale.
Alcune proteine sono ancorate alla
membrana da legami covalenti con le
catene idrocarburiche degli acidi
grassi
(a)
Thy-1 protein
Cys
Mannose
1
o=C
Glycosylphosphatidylinositol
NH2
Mannose
CH2
I
O
CH2-0-P-O- Mannose
1
O
N-acetylgalactosamine
N-acetylglucosamine
0
0-P-
Inositol
H2C-0
H2C-CH
-0+
0
Exoplasmic face
(H2C)12 (CH2)12
H3C
-CH3
CH3 CH3
HC
CCH
HC
C-CH3
HC
ECH
HCF
C-CH3
CH3
CH3
HC
Palmitate
Myristate
CH2
(CH2)14
(CH2)12
C=0
C=0
Cytosolic
face
0
S
NH
1
1
H3C-0-C-Cys-Lys-Cys
Gly
Ser
Ras protein
v-Src protein
(b)
+0-0-
(c)
-
Mannose -0-P-O-CH2-CH2-NH
O=L-0
0
-
Farnesyl
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
