Biochimica: Metabolismo energetico, degradazione macromolecole e fosforilazione
Documento di Biochimica sulla lezione del 27/05/2020, focalizzato sul metabolismo energetico. Il Pdf descrive la degradazione delle macromolecole, la fosforilazione ossidativa e il ciclo di Krebs, con diagrammi esplicativi per studenti universitari di Biologia.
Mostra di più8 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Metabolismo Energetico e Degradazione delle Macromolecole
Continuiamo il metabolismo energetico, parlando di degradazione delle macromolecole per recuperare energia chimica (ATP). Abbiamo di fatto seguito il glucosio nella via glicolitica che diventa acido piruvico e adesso vediamo come quest'ultimo può essere trasformato in acetil coA e poi vediamo come degradare gli acidi grassi attraverso una beta-ossidazione.
Formazione di Acetil CoA dall'Acido Piruvico
L'acido piruvico perde il gruppo coo- sotto forma di co2 e il pacchetto energetico che si sviluppa dalla perdita di ciò lo troviamo nel legame tioestere. L'acetil coA è un composto con un legame tioestereo con delta g di idrolisi più alto di quello dell'auto per questo la rottura di questo legame fornisce energia per la formazione processi sintetici.
Beta-Ossidazione degli Acidi Grassi
L'altra grande via metabolica che produce l'acetil coA attraverso una beta-ossidazione che degrada gli acidi grassi. Quest'ultimi in un momento di benessere metabolico sono impacchettati nei trigliceridi di deposito. Il glucagone è il segnale di bassa concentrazione di glucosio ematico. Ciò induce la degradazione dei trigliceridi di acidi grassi affinché si possa formare acetil-coA e di conseguenza ATP. La lipolisi periferica contribuisce in maniera indiretta al mantenimento della glicemia. Dalla degradazione di un acido grasso si ottengono 106 molecole di ATP. Tutte le cellule che hanno i mitocondri e sono in presenza di ossigeno, dovendo scegliere tra poche molecole di ATP (glicolisi) e una marea di acidi grassi che formano una marea di ATP (ciclo di Krebs) scelgono gli acidi grassi e lasciano il glucosio che verrà messo a disposizione delle cellule che non possono scegliere gli acidi grassi (non hanno i mitocondri o non permettono il passaggio dei grassi),come ad esempio le cellule nervose.
Meccanismo d'Azione del Glucagone
Come agisce il glucagone? Percorso di trasduzione del segnale, si attiva la lipasi,ormone sensibile,che taglia i diacilgliceroli e libera le due molecole arrivando al fegato liberando glucosio mentre gli acidi grassi raggiungono i distretti tissutali delle albumine.
Processo di Beta-Ossidazione
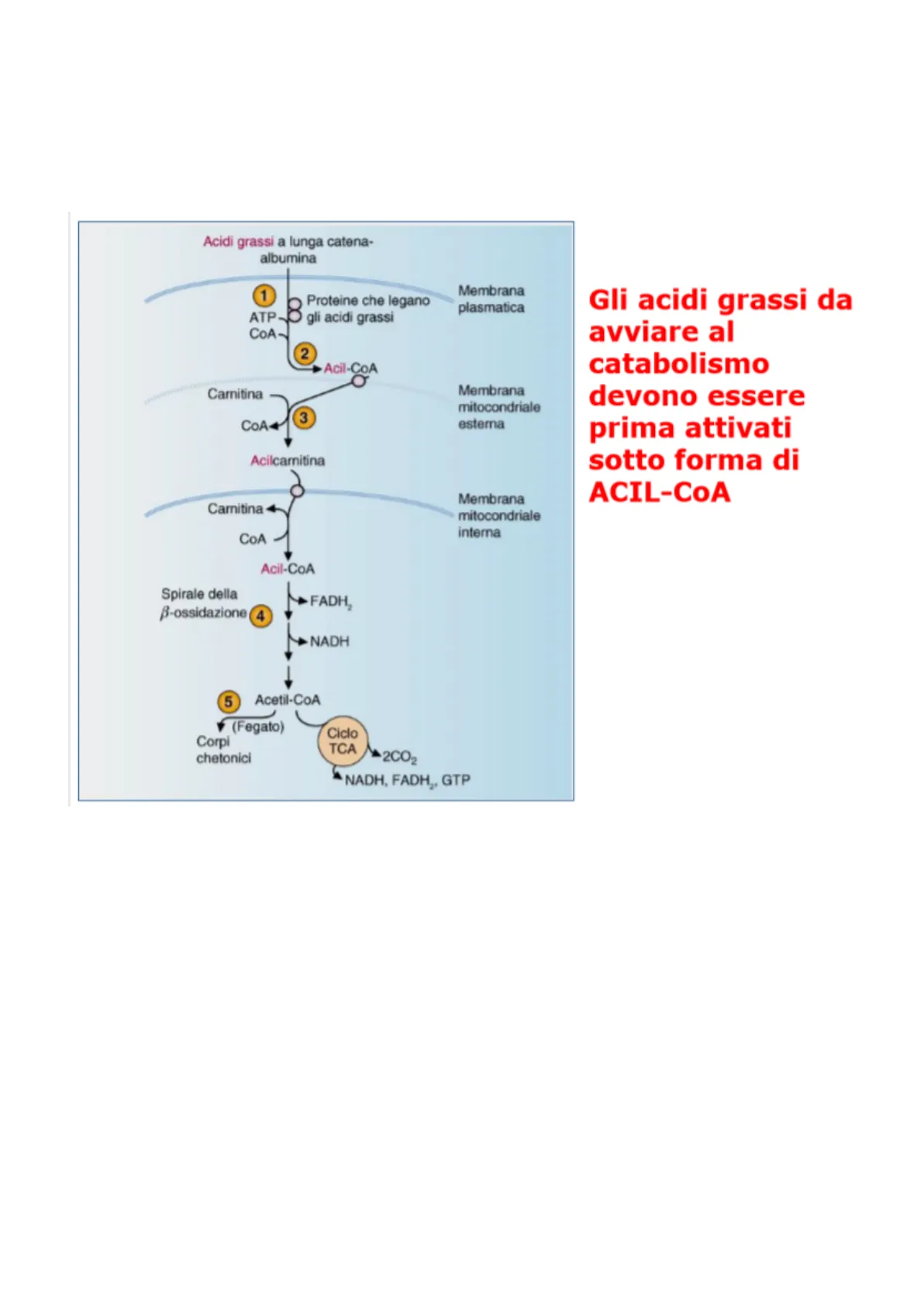
Gli acidi grassi vanno incontro al processo della beta-ossidazione,processo che consiste nel distacco di catene carboniose sotto forma di acetil coA. Ovviamente gli acidi grassi che entrano nella cellula devono essere attivati . Utilizzando l'idrolisi dell'ATP, viene legato al COO il coA con un legame ad alta energia di idrolisi.Questa attivazione avviene nel citosol. Entra nel mitocondrio attraverso il sitea della acilcarnitina. Sistema di antifurto che consente il passaggio dell'acido grasso rilasciando l'acil coA. A quel punto prende la via della beta ossidazione.
Acidi grassi a lunga catena- albumina 1 8 Proteine che legano gli acidi grassi Membrana plasmatica ATP CoA- 2 Acil-CoA Carnitina Membrana mitocondriale esterna COA3 Acilcarnitina Carnitina Membrana mitocondriale interna CoA Acil-CoA Spirale della B-ossidazione FADH, 4 NADH 1 5 Acetil-CoA V (Fegato) Ciclo TCA Corpi chetonici +2CO2 NADH, FADH2, GTP Gli acidi grassi da avviare al catabolismo devono essere prima attivati sotto forma di ACIL-COA
L'acido palmitico lega al gruppo COO l'acetil coA.Ora dobbiamo far uscire l'unità carboniose sotto forma di acetil-coA, per fare ciò dovremmo stacca il legame tra i carboni alfa e beta ma non esiste enzima che lo possa fare perché è troppo forte dunque ossidiamo il carbonio beta a C=O. Grazie a questo ponte questo legame tra alfa e beta si labilizza e esiste l'enzima che riesce a tagliare. Si chiama beta ossidazione perché è un processo a tappe che consente di realizzare il legame tra carbonio alfa e beta in maniera tale da poter rompere l'unità bicarboniosa sotto forma di acetil coA.
Ossidazione del Carbonio e Tappe
Come si ossida il carbonio? In 3 tappe diverse. Interviene una prima deidrogenasi FAD dipendente. Il primo passaggio è una deidrogenazione del legame CH2-CH2 e si forma l'enoyl, composto di doppio legame carbonio carbonio. Poi avviene un idratazione. Si forma l'idrossiacyl.Avviene una seconda deidrogenazione NAD dipendente formando il doppio legame C=O. Cioè il beta chetoacil. A questo punto arriva l'enzima tiolasi che può rompere il legame. Avviene l'eliminazione dell'acetil coA e quello che era acido palmitico è rimasto pari ma ha perso due atomi di carbonio. Diventa a 14 atomi e rifà tutti gli stessi passaggi per staccare la seconda molecola di acetil coA. Avviene questa sequenza di reazioni per 7 volte. Alla 6 volta avremo il beta chetoacil a 4 atomi di carbonio. Dunque nell'ultimo ciclo di reazioni si liberano 2 molecole di acetil coA.
Prodotti della Reazione di Beta-Ossidazione
PRODOTTI DI REAZIONE :
- 8 acetil-coA
- 7 FADH2 (1,5)
- 7 NADH (2,5)
Per ogni ciclo produciamo, indirettamente dalle deidrogenasi, 4 molecole di ATP dunque 28 in totale alla fine delle 7 ripetizioni. Le 8 molecole di acetil coA adesso dovranno entrare nel ciclo di Krebs per formare altro ATP.
Spirale Degradativa della Beta-Ossidazione
LA B-OSSIDAZIONE E' UNA SPIRALE DEGRADATIVA
C14 Acetyl -CoA C12 Acetyl -CoA C10 Acetyl -CoA C8 Acetyl -CoA C6 Acetyl -CoA Ciclo dell'acido citrico + FOXO C4 Acetyl -CoA Acetyl -CoALa formazione di acetil-coA è solo mitocondriale. Dobbiamo capire in che modo si ottengono 106 molecole di ATP.
Ciclo di Krebs
A questo punto nel ciclo di Krebs entrato acetil-coA che provengono dal l'acido piruvico e dalla beta-ossidazione. L'acetil-coA entrando nel ciclo di Krebs libererà CO2 e produrremo una molecola di ATP e ancora quote di coenzimi ridotti che poi arriveranno dove dovranno arrivare per essere ossidati e liberare ATP.
Fase 2 del Catabolismo
Fase 2 del catabolismo Ciclo di Krebs (4 tappe che rilasciano elettroni). Il substrato del ciclo di Krebs è acetilcoA che si condensa sotto forma di acido ossalacetico (chetoacido dicarbossilico). Ha due gruppi COO- a 4 atomi di carbonio. In realtà è il chetoacido di un amminoacido (l'acido aspartico). Quindi, da un lato il substrato è l'acetil coA perché il ciclo di krebs parte dalla condensazione di questo con l'acido ossalacetico con un enzima particolare (citrato sintasi) che porta alla formazione del citrato. L'acido citrico subisce una prima decarbossilazione ossidativa, si forma l'alfa che togu carat. Subisce una seconda decarbossilazione, si forma l'alfa succinil coA che è ancora ad un legame tioestere. Tanta alta energia che può essere usata per fosforilare un GDP a GTP. L'acido subisce un'altra decarbossilazione che forma l'acido fumarico, il quale viene idratato a malato e poi a ossalacetato. (Quindi l'ossalacetato non è totalmente substrato ma alla fine lo otteniamo anche come prodotto). Tutti i coenzimi ridotti andranno a finire nel processo di ossidazione per ottenere di nuovo i due coenzimi ossidati.
Regolazione del Ciclo di Krebs
L'acido citrico esce dal mitocondrio per andare ad inibire la fosfochinasi 1. Come esce dal mitocondrio? Il ciclo di Krebs in un momento di benessere energetico rallenta a valle e ciò fa sì che aumenti la concentrazione di acido citrico. Elevate concentrazioni di acido citrico porteranno alla sua uscita dovuto al fatto che c'è un alto rapporto NAD NADH. Se siamo in carenza la cellula ha acidi grassi? Si Quando la cellula è in carenza energetica degrada i trigliceridi di deposito. L'acido ossalacetico in un momento di carenza viene usato per formare la gluconeogenesi. Quindi quando diciamo che si accumula citrato è perché la cellula è in uno stato di benessere energetico. In un momento di carenza metabolica non si avrà l'acido ossalacetico perché starà facendo metabolismo. Già con la beta ossidazione degli acidi grassi la cellula può sopportare la bassa concentrazione di ATP e far deviare l'acido ossalacetico verso la gluconeogenesi per rispondere alla richiesta di glucosio che hanno le cellule obbligate.Abbiamo detto che il citrato si accumula solo in una stato di benessere energetico ciò, però, non significa che in condizioni di malessere energetico il citrato non si formi, solo che se ne forma poco tanto da non potersi accumulare. Il citrato come abbiamo detto inibisce la fosforilasi-1 battistrada della via glicolitica poiché è presente tanto NADH per cui non c'è bisogno di formare altro acido piruvico dunque nel metabolismo basale della cellula non avrebbe senso formare altro acido piruvico.
Prodotti del Ciclo di Krebs
PRODOTTI CICLO DI KREBS:
- 1 FADH2
- 3 NADH (ridotto)
- 1 ATP.
10 molecole di ATP diventano 20 perché dalla molecola di glucosio si formano 2 molecole di acetil coA. Alle 28 molecole dobbiamo sommare tutte le molecole di ossidazione ottenute dall'ossidazione dell'acetil coA. Dalle 108 togliamo 2 per ottenere 106 per il fatto che l'acido grasso deve essere attivato aggiungendo acil-coA.
Stechiometria della Riduzione dei Coenzimi e Formazione di ATP
table 16-1 Stoichiometry of Coenzyme Reduction and ATP Formation in the Aerobic Oxidation of Glucose via Glycolysis, the Pyruvate Dehydrogenase Reaction, the Citric Acid Cycle, and Oxidative Phosphorylation
| Reaction | Number of ATP or reduced coenzymes directly formed | Number of ATP ultimately formed* |
| Glucose -> glucose 6-phosphate | -1 ATP | -1 |
| Fructose 6-phosphate -> fructose 1,6-bisphosphate | -1 ATP | -1 |
| 2 Glyceraldehyde 3-phosphate -> 2 1,3-bisphosphoglycerate | 2 NADH | 3-5 |
| 2 1,3-Bisphosphoglycerate -> 2 3-phosphoglycerate | 2 ATP | 2 |
| 2 Phosphoenolpyruvate -> 2 pyruvate | 2 ATP | 2 |
| 2 Pyruvate -> 2 acetyl-CoA | 2 NADH | 5 |
| 2 Isocitrate -> 2 @-ketoglutarate | 2 NADH | 5 |
| 2 a-Ketoglutarate -> 2 succinyl-CoA | 2 NADH | 5 |
| 2 Succinyl-CoA -> 2 succinate | 2 ATP (or 2 GTP) | 2 |
| 2 Succinate -> 2 fumarate | 2 FADH2 | 3 |
| 2 Malate -> 2 oxaloacetate | 2 NADH | 5 |
| Total | 30-32 |
*This is calculated as 2.5 ATP per NADH and 1.5 ATP per FADH2. A negative value indicates consumption.
Ciclo di Krebs: Via Anfibolica
Abbiamo visto il ciclo di Krebs come una via degradativa. Di conseguenza contestualizzato in un contesto catabolico. In realtà il ciclo di Krebs è una via anfibolica perché molti suoi intermedi di reazione intermedi sono precursori i vie biosintetiche. Se siamo in un momento di carenza energetica degradiamo l'acido glutammico in alfa chetoglutarico potendo così fare i processi inversi. In momento di benessere energetico il citrato esce e giunge nel citosol. Nel citosol se la cellula non necessita nulla il citrato, come abbiamo già detto, va a inibire la via glicolitica se invece la cellula è in carenza il citrato subisce una reazione attraverso cui viene scisso in acido ossalacetico e acetil coA, perché solo attraverso il citrato possiamo mandare acetil coA nel citosol. Una volta avvenuta questa scissione avviene la sintesi di acido glutammico e colesterolo. Detto questo, perché se dobbiamo scindere l' acido citrico in ossalacetato e acetil coA serve ATP? Perché serve a formare il legame tioestereo tra l'acetil e il coA. L'ATP serve per la scissione del citrato nel caso della sintesi degli acidi grassi non serve. Il citrato è il punto intermedio indiretto della sintesi degli acidi grassi.
Reazioni Anaplerotiche
REAZIONI ANAPLEROTICHE Se da una parte abbiamo intermedi che sfuggono al ciclo di Krebs dall'altra abbiamo reazioni di riempimento cioè anaplerotiche. Reazioni differenti e sono punti di entrata.
Tabella delle Reazioni Anaplerotiche
table 16-2 Anaplerotic Reactions
| Reaction | Tissue(s)/organism(s) |
| Pyruvate + HCO3 + ATP oxaloacetate + ADP + Pi | Liver, kidney |
| PEP carboxykinase | |
| Phosphoenolpyruvate + CO2 + GDP oxaloacetate + GTP | Heart, skeletal muscle |
| essere | |
| Phosphoenolpyruvate + HCO3 oxaloacetate + P. | Higher plants, yeast, bacteria |
| trasformato | |
| acido | |
| ossalacetico. | |
| in | |
| malie enzyme | |
| Pyruvate + HCO3 + NAD(P)H malate + NAD(P)* | Widely distributed in eukaryotes and prokaryotes |
COOH 1 ATP + HCO3 + C=O I CH3 Piruvato Piruvato Biotina carbossilasi Acetil-CoA COOH - C=0 + ADP + P. 1 CH2 COO" Ossalacetato Il ciclo di Krebs è regolato con i metaboliti che sono alla base del benessere o malessere energetico. pyruvate carboxylase Un esempio è l'acido piruvico che può entrare nel ciclo di Krebs perché può PEP carboxylase
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
