La neurotrasmissione: meccanismi sinaptici e malattia di Parkinson
Documento sulla neurotrasmissione. Il Pdf, utile per lo studio universitario di Biologia, esplora i meccanismi sinaptici, la classificazione dei neurotrasmettitori e la malattia di Parkinson, analizzandone eziopatologia, patofisiologia e trattamenti farmacologici.
Mostra di più11 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
La Neurotrasmissione
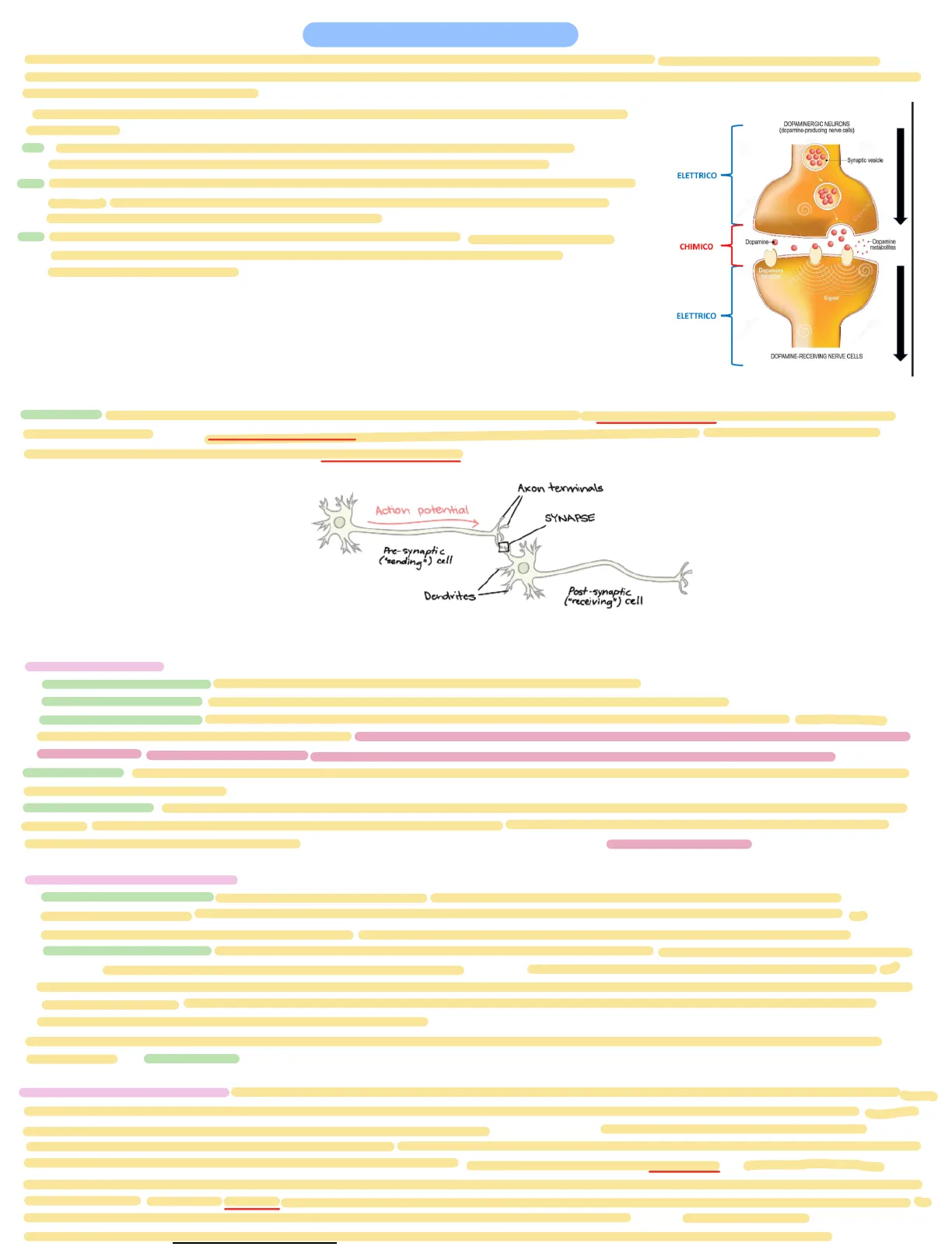
Il trasferimento dell'informazione da un neurone a un altro è chiamato trasmissione sinaptica. Nella maggior parte delle sinapsi, l'informazione viaggiando lungo l'assone nella forma di impulsi elettrici viene convertita nella parte terminale in un segnale chimico che attraversa lo spazio Inter sinaptica.
La trasmissione dell'informazione a livello neurale avviene attraverso meccanismi di natura neurochimica:
- Il segnale elettrico prodotto dal neurone presinaptico viaggia sino al terminale dell'assone dopo viene convertito in segnale chimico a livello della sinapsi.
- Nella membrana post sinaptica, questo segnale chimico viene riconvertito in un segnale elettrico. Il segnale chimico e chiamato neurotrasmettitore e viene immagazzinato e rilasciato dalle vescicole sinaptiche nel terminale.
- Diversi neurotrasmettitori sono usati da diversi tipi di neuroni. Questa trasformazione dell'informazione elettrica-chimica-elettrica rende possibili molte delle abilità computazionali del cervello.
H DOPAMINERGIC NEURONS (dopamine-producing nerve cells) Synaptic vesicle ELETTRICO dreamsti CHIMICO Dopamine- +Dopamine metabolites Dopamine receptor Signal ELETTRICO DOPAMINE-RECEIVING NERVE CELLS
La Sinapsi
La sinapsi: consta di due parti ovvero la parte presinaptica e quella postsinaptica. La zona presinaptica possiede generalmente un terminale assonico, mentre la parte postsinaptica può essere il dendrite o il soma di un altro neurone. Lo spazio tra la membrana presinaptica e postsinaptica viene chiamato spazio intersinaptico (20 mm).
Axon terminals Action potential SYNAPSE Pre-synaptic (" sending") cell Dendrites Post-synaptic ("receiving") cell
Tipi di Sinapsi
- Sinapsi asso-dendritiche: il bottone sinaptico termina sulla spina dendritica di un dendrite;
- Sinapsi asso-somatiche: il bottone sinaptico termina direttamente sul soma del neurone post-sinaptico;
- Sinapsi asso-assoniche: il bottone sinaptico termina sulla parte terminale dell'assone del neurone post-sinaptico. Questo tipo di sinapsi permette una facilitazione presinaptica: un neurone presinaptico A è in grado di influenzare una singola sinapsi del neurone presinaptico B, anziché l'intero neurone, e va ad agire sul neurone postsinaptico C, facilitandone o inibendone l'azione.
Sinapsi Dirette e Non Dirette
Sinapsi dirette: una sinapsi è diretta quando il neurotrasmettitore viene rilasciato dal bottone presinaptico in stretta prossimità al sito nel quale si trovano i recettori.
Sinapsi non dirette: una sinapsi è non diretta quando il sito di rilascio del neurotrasmettitore è situato a una certa distanza dal sito di ricezione. In questo caso l'assone presenta solitamente delle varicosità, cioè dei rigonfiamenti nei quali sono contenute numerose vescicole contenenti il neurotrasmettitore. Proprio per questo vengono anche chiamate sinapsi a filo di perle.
Tipi di Neurotrasmettitori
- Piccoli neurotrasmettitori: sono di diversa natura chimica. Vengono sintetizzati direttamente nel citoplasma a livello della terminazione sinaptica; vengono immagazzinati in piccole vescicole grazie all'apparato di Golgi che si trova nel terminale; le vescicole che possiedono il neurotrasmettitore, vengono immagazzinate in gruppi in prossimità della membrana sinaptica.
- Grandi neurotrasmettitori: sono di natura peptidica e sono catene composte da 3-36 residui, per cui sono considerati neuropeptidi. Vengono sintetizzati nel citoplasma del soma grazie ai ribosomi; vengono immagazzinati all'interno di vescicole grazio al Golgi; le vescicole che possiedono il neuropeptide, vengono trasportate lungo i microtubuli dal soma al terminale sinaptico a una velocità di circa 40cm al giorno. Questo tipo di vescicole hanno dimensioni maggiori rispetto a quelle dei piccoli neurotrasmettitori e non formano aggregati in prossimità della membrana sinaptica.
Molte specie neuronali sono in grado di sintetizzare e liberare più neurotrasmettitori, di solito un piccolo neurotrasmettitore e un neuropeptide -> coesistenza.
Rilascio dei Neurotrasmettitori
Rilascio dei neurotrasmettitori: il rilascio del neurotrasmettitore è attivato dall'arrivo di un potenziale d'azione nel terminale assonico. La depolarizzazione della membrana del terminale induce l'apertura dei canali-voltaggio dipendenti per il calcio nelle zone attive. Sapendo che la concentrazione ionica interna del Ca2+ a riposo è molto bassa, di conseguenza si riverserà nel citoplasma del terminale assonico per tutto il tempo che il canale rimarrà aperto. L'innalzamento quindi della concentrazione del calcio è il segnale che induce il rilascio del neurotrasmettitore da parte delle vescicole sinaptiche. Questo rilascio avviene per esocitosi -> la membrana della vescicola sinaptica si fonde con la membrana presinaptica nella zona attiva permettendo al contenuto della vescicola di riversarsi nella fessura sinaptica. Le proteine SNARE sono una grande famiglia di proteine composta da più di 60 membri nelle cellule dei mammiferi. Il loro ruolo è proprio quello di regolare la fusione delle vescicole con la membrana bersaglio, quindi mediano l'esocitosi.
Per quanto riguarda i piccoli neurotrasmettitori, l'ingresso degli ioni Ca porterà sempre al rilascio di una certa quantità dineurotrasmettitore per cui si parla di rilascio pulsatile.
Il rilascio dei neuropeptidi invece avviene in modo più stabile e graduale solo se il citoplasma della terminazione ha raggiunto una certa concentrazione di ioni Ca e ciò avviene in seguito all'attivazione sostenuta del neurone.
I Recettori
I neurotrasmettitori si legano con specifiche molecole chiamate recettori, per questo prendono anche il nome di ligandi. Un neurotrasmettitore può legarsi a diversi tipi di recettore -> sottotipi recettoriali. Ognuno di questi sottotipi di recettore riesce a indurre nel neurone nel quale è localizzato, uno specifico effetto stimolando o inibendo. Il legame di un neurotrasmettitore a uno dei suoi sottotipi recettoriali può influenzare il neurone postsinaptico in uno dei due modi fondamentali, a seconda che il recettore sia ionotropico (associ8ati a canali ionici girando-dipendenti) o metabotropico (associati a proteine di segnalazione e proteine G sensibili alla guanosina trifosfato).
Recettori Ionotropici
Recettori ionotropici: quando un neurotrasmettitore si lega a uno di questi recettori, il canale ionico associato si apre o si chiude immediatamente, portando a un potenziale postsinaptico. In alcuni casi si produce una depolarizzazione transitoria che viene chiamata PPSE (potenziale postsinaptico eccitatorio) perché il neurotrasmettitore determina l'apertura dei canali del sodio e quindi l'aumento di ioni sodio nel neurone; invece, si crea una PPSI (potenziale postsinaptico inibitorio) cioè l'iperpolarizzazione transitoria dovuta all'apertura dei canali del K+ o del CI- da parte del neurotrasmettitore da cui consegue rispettivamente un aumento del flusso di ioni K+ verso l'esterno del neurone o flusso di ioni CI- verso l'interno, rendendo l'interno del neurone ancora più negativo.
Generazione di PPSE
Generazione di un PPSE: un impulso che arriva nel terminale presinaptico induce il rilascio del neurotrasmettitore. Le molecole di quest'ultimo si legano ai recettori che si trovano nei canali ionici tramsettitore-dipendenti nella membrana postsinaptico. Se i canali si aprono per far entrare gli ioni sodio, allora la membrana si depolarizza -> questo è il caso dei recettori nicotinici per l'acetilcolina che troviamo nella placca neuromuscolare e la cui attivazione permette la contrazione dei muscoli.
Generazione di PPSI
Generazione di un PPSI: se i canali trasmettitore-dipendenti sono permeabili al Cl- allora avverrà una iperpolarizzazione della cellula. Questo è il caso dei recettori GABA A per il GABA, che è il principale neurotrasmettitore inibitorio del SNC. Avviene una inibizione proprio perché il potenziale viene portato a valori lontani da quelli di riposo.
Recettori Metabotropici
Recettori metaboprotici: gli effetti di questi recettori sono più lenti e vari. Esistono diversi tipi di questi recettori e ciascuno di essi è collegato a una proteina di segnalazione, ovvero una proteina transmembrana (che attraversano 7 volte). Il recettore si attacca a una porzione della proteina sul lato extracellulare, mentre sul versante interno del neurone è situata una proteina G. Quando il neurotrasmettitore si lega al recettore metabotropico, una subunità della proteina g si dissocia e si possono prendere due vie:
- La subunità della proteina g coinvolta si lega a un canale ionico inducendo un PPSE o un PPSI;
- Oppure può stimolare la sintesi o l'attività di una sostanza chimica chiamata secondo messaggero, questo si diffonde nel
citoplasma e può influenzare l'attività del neurone in diversi modi:
- Entrare nel nucleo e influenzare l'attività del DNA, quindi influenzare l'espressione genica, in questo caso il recettore metabotropico può avere effetti radicali di lunga durata;
- o Può dare vita a cascate enzimatiche che porteranno a modificazioni più o meno durature dell'attività del neurone.
Autorecettori
Autorecettori I recettori dei neurotrasmettitori si possono trovare anche nella membrana del terminale assonico presinaptico. I recettori presinaptici sensibili al neurotrasmettitore rilasciato dalla stessa terminazione presinaptica sono detti autorecettori. Di solito questi recettori sono accoppiati alla proteina G e promuovono la formazione di secondi messaggeri. Ci sono molteplici conseguenze legate alla loro attivazione, ma un effetto comune è l'inibizione del rilascio del neurotrasmettitore e in alcuni casi della sua stessa sintesi. Ciò permette al terminale presinaptico di autoregolarsi. Gli autorecettori sembra che lavorino come delle valvole di sicurezza per ridurre la liberazione del neurotrasmettitore quando la sua concentrazione nei pressi del terminale presinaptico raggiunge livelli troppo alti, o al contrario stimolarla quando invece i suoi livelli di concentrazione sono troppo bassi.
Differenze tra Neurotrasmettitori
A questo punto, date le conoscenze, possiamo capire altre differenze tra piccoli neurotrasmettitori e neuropeptidi:
- I piccoli neurotrasmettitori vengono rilasciati solitamente a livello di sinapsi dirette e in genere attivano recettori ionotropi o metabotropi che agiscono sui canali ionici; sembra che questi neurotrasmettitori abbiano la funzione di trasmettere brevi e rapidi segnali eccitatori o inibitori a cellule adiacenti.
- I neuropeptidi tendono a essere rilasciati diffusamente e quasi tutti si legano a recettori metabotropici che agiscono tramite secondi messaggeri; la funzione dei neuropeptidi sembra essere quella di trasmettere segnali lenti, diffusi e di lunga durata, anche attraverso il coinvolgimento di modificazioni dell'espressione genica.
Meccanismi di Inattivazione
Senza un meccanismo di inattivazione del neurotrasmettitore, questo rimarrebbe continuamente nella sinapsi stimolando i recettori e questo porterebbe a un blocco della trasmissione sinaptica -> per questo esistono meccanismi che regolano questa inattivazione del neurotrasmettitore una volta che ha esercitato i suoi effetti attraverso l'interazione con il recettore. I principali meccanismi di inattivazione sono:
- Ricaptazione (ricattura nel terminale presinaptico): è il metodo più comune per bloccare l'attività del neurotrasmettitore nella sinapsi. Gran parte dei neurotrasmettitori, una volta rilasciati nella sinapsi, viene riassorbita quasi immediatamente all'interno dei terminali presinaptici tramite un meccanismo attivo mediato da trasportatori; quindi il neurotrasmettitore viene agganciato dal trasportatore e portato attivamente all'interno del terminale presinaptico che l'aveva liberato. Alcune molecole bloccano questi trasportatori, ad esempio la cocaina blocca la ricattura della dopamina, mentre degli antidepressivi noti con la sigla SSRI, bloccano la ricattura della serotonina.
- Degradazione enzimatica nella fessura sinaptica: inattiva il neurotrasmettitore a livello della sinapsi grazie a specifici enzimi. Ad esempio l'acetilcolina è uno di quei trasmettitori per cui la degradazione enzimatica rappresenta il meccanismo principale di inattivazione che avviene grazie all'enzima acetilcolinesterasi.
In generale, il terminale sinaptico è molto efficiente, in quanto alcuni dei processi che vi avvengono servono a riutilizzare e/o riciclare molte delle molecole che partecipano alla sua attività; ciò succede, ad esempio, con i neurotrasmettitori, parte dei quali una volta ricatturati vengono di nuovo immagazzinati all'interno delle vescicole che a loro volta vengono anche riutilizzate come magazzini per il neurotrasmettitore.
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
