Istologia ed Embriologia: sviluppo del mesoderma e formazione dei somiti
Documento di Università su Istologia ed Embriologia – Lezione 30. Il Pdf esplora lo sviluppo del mesoderma e la formazione dei somiti, con dettagli sui derivati del foglietto mesodermico e l'influenza dei geni HOX, utile per lo studio della Biologia.
Mostra di più20 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Istologia ed Embriologia - Lezione 30
Sbobinatore: Adriano Caggia Docente: Giuseppe Bonaventura Data: 21/12/2023
INDICE
- Derivati del foglietto mesodermico
- Mesoderma parassiale
- Somiti
- Geni omeotici hox
- Scoperta delle trasformazioni omeotiche
- Sinpolidattilia
- Evoluzione dei somiti
- Segnali molecolari che influenzano la differenziazione dei somiti
- Mesoderma intermedio
- Mesoderma laterale
- Regolazione dell'induzione neurale
- I ripiegamenti trasversali e longitudinali dell'embrione
- Annessi amnio-fetali
- La placenta
- Membrana o barriera placentare
- Funzioni della placenta
- Difetti placentari
- Cordone ombelicale
DERIVATI DEL FOGLIETTO MESODERMICO
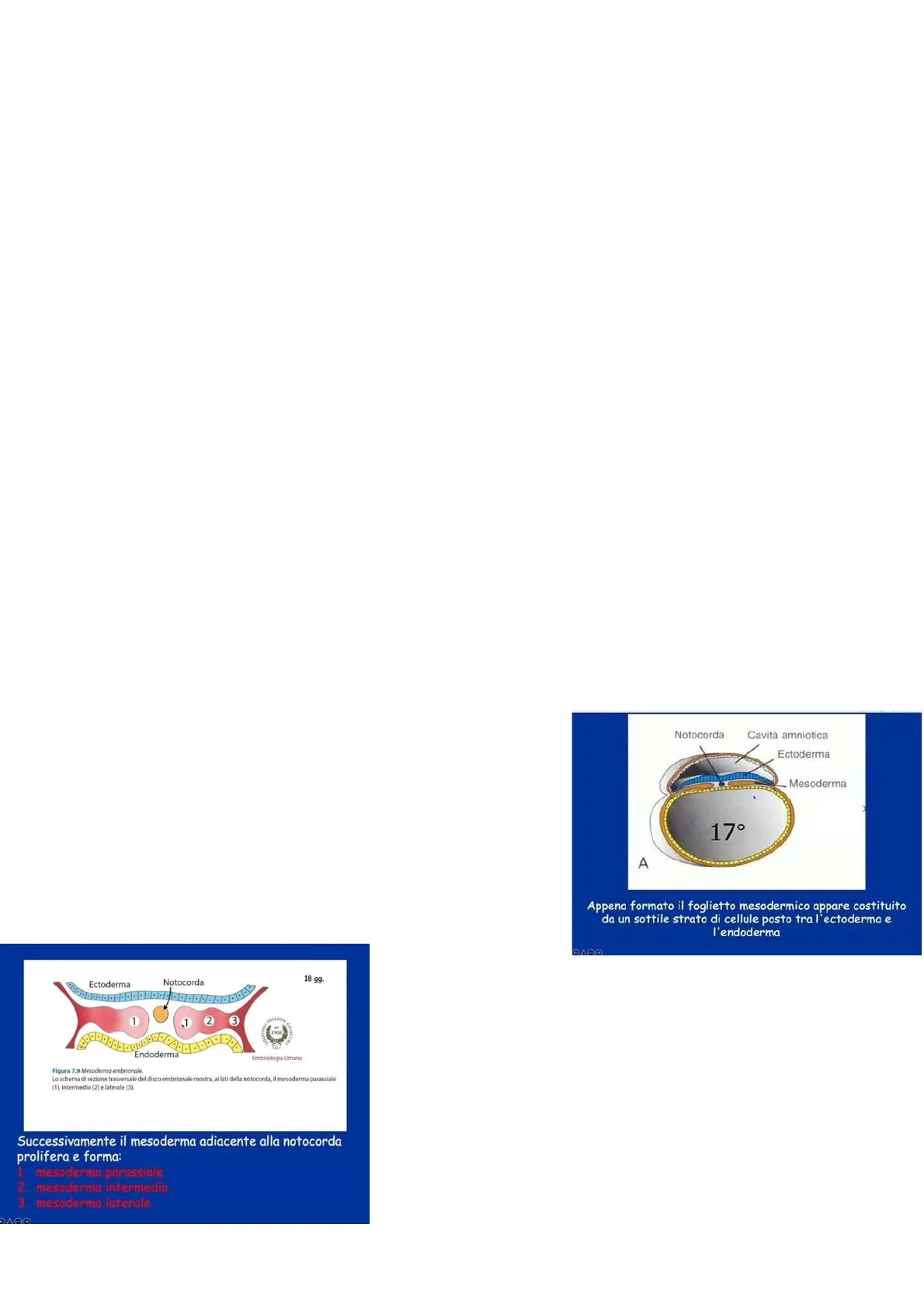
Al 17 esimo giorno (quindi a fine gastrulazione) il mesoderma, osservato in una sezione trasversa dell'embrione, appare costituito da un sottile strato di cellule che si va ad interporre tra ectoderma ed endoderma. Come si osserva dal disegno due lamine cellulari sono disposte ai lati della notocorda, questo perché si sta osservando la sezione di una regione craniale più cefalica dell'embrione dove si trova la notocorda. Se ci si fosse trovati in una regione più posteriore si sarebbe vista una lamina unica non separata dalla notocorda.
Notocorda Cavità amniotica Ectoderma Mesoderma 17° A Appena formato il foglietto mesodermico appare costituito da un sottile strato di cellule posto tra l'ectoderma e l'endoderma DADO Ectoderma Notocorda 18 gg 1 1 2 3 1908 Endoderma Embriologia Umana Figura 7.9 Mesoderma embrionale. Lo schema di sezione trasversale del disco embrionale mostra, ai lati della notocorda, il mesoderma parassiale (1), intermedio (2) e laterale (3).
Successivamente il mesoderma adiacente alla notocorda prolifera e forma:
- mesoderma parassiale
- mesoderma intermedio
- mesoderma laterale
Successivamente, dal 18 esimo giorno, a partire dalla regione più cefalica dell'embrione, il mesoderma, posizionato accanto alla notocorda, inizia a proliferare formando due grossi cordoni, che vanno a costituire il mesoderma parassiale (n1).
Questo è quanto si può osservare in una sezione trasversa, mentre in una sezione longitudinale si vedrebbero effettivamente i due cordoni come descritti.
1Il mesoderma parassiale è in continuazione, lateralmente, con dell'altro mesoderma meno spesso distinto in: - intermedio (n2) - laterale (n3) (abbiamo parlato di queste 3 diverse sezioni del mesoderma quando abbiamo visto che c'è una predestinazione tra le diverse cellule che si invaginano nelle varie regioni del solco primitivo e nella fessura primitiva). Per riassumere: al 18 esimo giorno, il mesoderma ha già cominciato a diversificarsi in: parassiale, intermedio e laterale.
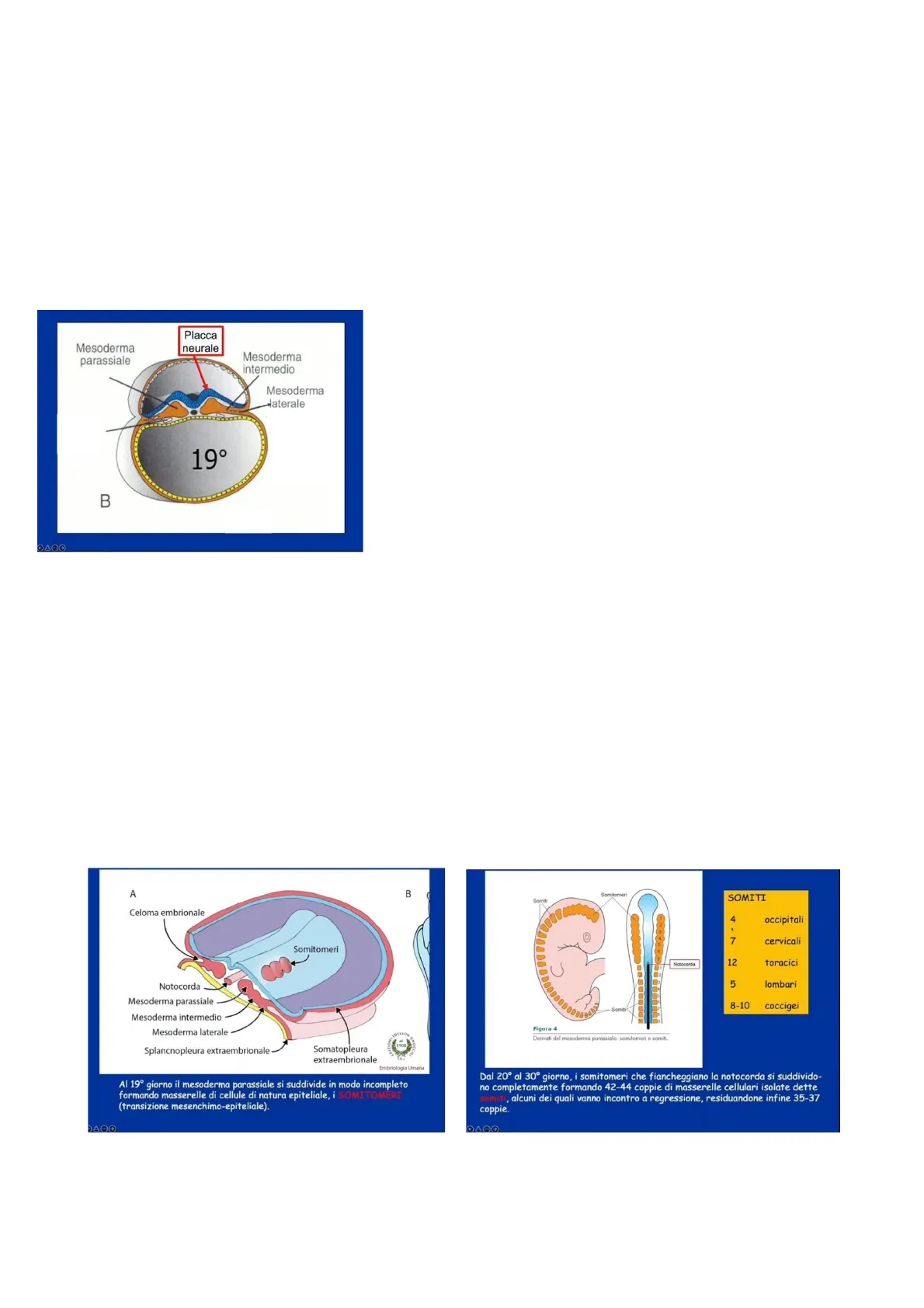
Placca neurale Mesoderma parassiale Mesoderma intermedio Mesoderma aterale B Questo disegno suggerisce che, già al 19 esimo giorno, quando stanno avvenendo delle modificazioni a carico del mesoderma, contemporaneamente si verifica un'altra modificazione sull'ectoderma: la formazione della placca neurale (questo conferma come gli eventi dal punto di vista temporale vengono a sovrapporsi).
MESODERMA PARASSIALE
Lo sviluppo del mesoderma continua e i due cordoni di cui si è già parlato intorno al 19 esimo giorno cominciano a segmentarsi in modo incompleto, cioè si dividono in piccoli ammassi cellulari ancora collegati tra loro. Questi ammassi cellulari sono di natura epiteliale e vengono chiamati somitomeri.
Questo processo di segmentazione comincia a partire dalla zona più cefalica dell'embrione e prosegue via via in senso caudale in direzione posteriore.
Quindi dal mesenchima che prolifera dal contesto del mesoderma parassiale si formano questi piccoli ammassi di cellule che diventano di natura epiteliale. Questo è un fenomeno tipico di transizione mesenchimo-epiteliale: trasformazione di cellule mesenchimali in epiteliali, processo inverso a quello precedentemente visto, durante il distacco delle cellule epiblastiche invaginate durante la gastrulazione.
A B Celoma embrionale Somitomeri Notocorda Mesoderma parassiale Mesoderma intermedio Mesoderma laterale Splancnopleura extraembrionale Somatopleura extraembrionale MOCCA Embriología Umana Al 19º giorno il mesoderma parassiale si suddivide in modo incompleto formando masserelle di cellule di natura epiteliale, i SOMITOMERI (transizione mesenchimo-epiteliale).
Somitomeri Somit 1 Notocorda Somiti Figura 4 Derivati del mesoderma parassiale: somitomeri e somiti.
SOMITI 4 occipitali + . 7 cervicali 12 toracici 5 lombari 8-10 coccigei Dal 20° al 30º giorno, i somitomeri che fiancheggiano la notocorda si suddivido- no completamente formando 42-44 coppie di masserelle cellulari isolate dette somiti, alcuni dei quali vanno incontro a regressione, residuandone infine 35-37 coppie.
SOMITI
Formazione dei somiti
Dal 20 esimo giorno e fino a fine mese (30 esimo giorno) i somitomeri che si sono via via andato formando, escludendo i primi 7, (quelli che si trovano in una posizione anteriore rispetto alla notocorda), andranno incontro ad un'ulteriore evoluzione: si separeranno tra loro, formando piccoli ammassi cellulari isolati e staccati gli uni dagli altri i così detti SOMITI.
Quindi i primi sette restano somitomeri, gli altri si trasformano in somiti.
Ripetendo: al 19 esimo giorno sono tutti somitomeri, al 20 esimo giorno inizia un altro processo di distaccamento dei somitomeri tra loro escludendo i 7 che si trovano in una posizione anteriore rispetto alla notocorda; gli altri, che costeggiano la notocorda, si andranno a staccare completamente costituendo i somiti.
Avremo alla fine 42-44 paia di somiti che sono distinti in:
- 4 occipitali;
- 7 cervicali;
- 12 toracici;
- 5 lombari;
- 8-10 coccigei;
1 dei 4 occipitali e 5-7 dei coccigei sono destinati alla degenerazione, e dunque alla fine il numero definitivo sarà di 35/37 coppie.
Questi somiti rappresentano strutture temporanee che però risultano fondamentali per l'organizzazione segmentale dell'embrione. Sono cioè necessari per la metameria o struttura metamerica dell'organismo. Essa riguarda l'organizzazione corporea di tutti i vertebrati. Quindi molto precocemente un organismo in via di sviluppo appare già suddiviso, considerando la sua lunghezza, in vari segmenti corporei che sono simili ma non uguali. In ciascuno di essi, è possibile osservare una certa ripetitività, una certa serialità. Si tratta di ripetizioni di componenti che prevedono le stesse unità funzionali.
Per fare un richiamo alla pratica clinica, a livello cutaneo ognuno di questi dermatomeri (segmenti corporei a livello cutaneo), risulterà innervato sia da una specifica radice nervosa motoria, sia anche da una radice nervosa sensitiva. Sono due radici nervose che nascono da un preciso metamero (tratto) del midollo spinale; quindi ognuno di questi dermatomeri, dipende da un preciso segmento del midollo spinale.
GENI OMEOTICI HOX
I SOMITI sono strutture transitorie ma determinanti per l'organizzazione segmentale dell'embrione (metameria).
Suddivisione dell'organismo (lungo l'asse antero- posteriore) in segmenti corporei seriali (metamer))
Distribuzione segmentale del miotomi in un feto di sei settimane
- ---
La specificazione dei diversi segmenti corporei è controllata dei cosiddetti geni omeotici HOX. preposti all'attivazione controllata di batterie di altri geni (PAX, SOX, OTX) adibiti allo sviluppo del piano architettonico dell'organismo che ne deriva sarà innervata da piùi di un singolo segmenin spinale GAGO I SOMITI sono strutture transitorie ma determinanti per l'organizzazione segmentale dell'embrione (metameria).
Distribuzione segmentale dei miotomi in un feto di sei settimane I miotomi, similmente al somiti da cui derivano, presentano una distribuzione seg mentale. Ciascun segmento è innervato da un paio di nervi spinali derivanti dal relativo segmento di midollo spinale. Dalla piccola porzione dorsale del miotomo origina l'epi mero; mentre la porzione ventrale più larga porta alla formazione dell'ipomero. Spesso, miotomi adiacenti convergono fra di loro; ne segue che la massa muscolare scheletrica che ne deriva sarà innervata da più di un singolo segmento spinale.
DAOO A determinare la specificità di ognuno di questi segmenti corporei troviamo dei geni speciali, sono i geni omeotici HOX. Tali geni sono in grado di svolgere un'azione di controllo di livello molto elevato, un controllo operato su una serie di altri geni che si rendono responsabili della realizzazione del piano architettonico dell'embrione e che stabiliscono la normale successione dei segmenti corporei.
SCOPERTA DELLE TRASFORMAZIONI OMEOTICHE
La scoperta delle trasformazioni omeotiche Nel 1894, l'inglese Bateson identificò negli artropodi delle varianti strutturali in cui una parte del corpo veniva sostituita da un'altra. Denominò queste "stranezze" trasformazioni omeotiche
Antennae Legs (a) Normal fly (b) Mutant fly Il termine "omeotico" deriva dallo studioso inglese Bateson, il quale si accorse, studiando gli artropòdi, che alcuni insetti mostravano strane mutazioni: avevano una parte del corpo a volte replicata, altre volte sostituita con una inutile. Per esempio si vide che in un segmento corporeo come la testa, a seguito di una di queste mutazioni, potevano spuntare delle zampe, o anche delle ali.
I geni HOX si è visto che codificano i relativi fattori di trascrizione: fattori HOX, caratterizzati da un elemento comune a tutti questi geni, cioè una specifica sequenza di nucleotidi di DNA detta omeobox, che appare simile in tutti gli eucarioti, vertebrati e invertebrati.
È stata scoperta nel 1984, all'interno dei geni Antennapedia e Bithorax della Drosophila Melanogaster. Questa sequenza consiste in un tratto di DNA di 180 basi, 60 triplette, dunque un peptide di 60 amminoacidi. Tale polipeptide si chiama dominio omeotico o omeodominio. Esso fa parte della struttura di tutti i fattori di trascrizione HOX e serve a permettere ad ognuno di questi fattori di legarsi al DNA. C'è anche una parte variabile in ogni fattore di trascrizione HOX, la quale varia, appunto, nella sequenza aminoacidica: è quella che poi si rende responsabile della funzione di controllo svolta da quel determinato fattore HOX. A livello molecolare questo omeodominio presenta una struttura particolare: consiste in tre alfa-eliche in continuazione tra di loro.
Nell'immagine, l'alfa-elica rossa (n3) è responsabile del contatto principale con DNA: alfa-elica di riconoscimento. Le altre due servono a stabilire altri contatti con specifici fattori di trascrizione, quindi legano fattori di trascrizione che in quella precisa sede riescono a far esprimere il DNA.
Geni omeofici e omeodominio I geni omeotici (HOX) codificano i fattori di trascrizione HOX e sono accomunati dalla presenza dell' omeobox, una sequenza di DNA di 180 bp molto conservata evolutivamente, individuata per la prima volta nel 1984 all'interno dei geni Antennapedia e Bithorax di Drosophila melanogaster.
La sequenza dell'omeobox (DNA) corrisponde all'omeodominio dei fattori di trascrizione HOX, una regione di 60 aminoacidi che lega il DNA, regolando l'espressione genica.
Oltre che dall'omeodominio (per il legame con il DNA), ciascun fattore HOX risulta costituito da una regione variabile, artefice della sua specifica azione di controllo.
L'omeodominio è un modulo di 60 amminoacidi L'omeodominio ha 3 a-eliche 1 Braccio N-terminale 10 Elica 1 20 Glu byn Any Pro Arg The Ala Phe Sor Ss Giu Gần Leu Aia Arg Liiii Lys Ang Gầu Phú Aun Ghy Ang Lým Ang Giy Ang Gìn Thị Tyr Thư Ang Tyr Gin Thị Leu Gig Leu Gau Lại Gia Pre His Phe Ang Ang Lyn Lys Ang The Ser tie Glu The AsnVal Arg Phe Ala Cầu Giu Lys Set Phe Leu Ala Le eliche 1 e 2 sono sopra il DNA Il braccio N-terminale entra nella scanalatura secondaria 30 40 Aon Ang Tyr Lou The Gia Arg Ang Ang Glu Glu Las Ser Ss Giu Leu Gly Leu Đàn Ang Tyr Liêu The Arg Arg Arg Arg lio Giu Be Aia Hạ Aia Lên Cys Lêu Au Glu Lys Pro The Ser Glu Glu le Leu Low Be Nia Giu Gin ine His Met 42 41 Elica 3 Asn Câu Aia Giản Tên Lực Bịo Tỉp Phần Gin Aan Lys Ary Ala Lys Be Lya Lys Ser Agm Thị Gia Arg Gin Be Lys Be Trp Phơi Gìn Bạn Ang Ang Met Lyo Trp Lys Lys Giu Aan Giu Lys Giu Vai Bio Arg Vie Tru Phe Cys Aun Ang Ang Găn Lys Giu Lýu Ang Ba Aan 1 L'elica 3 entra nella scanalatura principale In molti organismi questi fattori di trascrizione sono essenziali per specificare, il destino differenziativo delle cellule durante il differenziamento e lo sviluppo embrionale.
A livello molecolare, l'omeodominio possiede una struttura secondaria definita da tre alfa-eliche, di cui la nº 3 entra nella scanalatura principale del DNA.
La sequenza omeobox è uguale in tanti animali. I geni omeotici sono stati riconosciuti in moltissimi animali, vertebrati e invertebrati, compreso l'uomo. La constatazione che tutti i geni omeotici si trovano in organismi molto diversi tra di loro la si può considerare come una prova che tali geni sono molto antichi e che gran parte degli animali che si conoscono l'hanno potuta ereditare da un antenato comune molto antico.
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
