Sistemas de Endomembranas: Retículo Endoplasmático y Aparato de Golgi
Documento de la Universidad sobre Sistemas de Endomembranas. El Pdf explora el retículo endoplasmático rugoso y liso, y el aparato de Golgi, detallando sus funciones en la síntesis de proteínas y lípidos, la desintoxicación y el transporte intracelular en Biología.
Ver más24 páginas

Visualiza gratis el PDF completo
Regístrate para acceder al documento completo y transformarlo con la IA.
Vista previa
Sistemas de Endomembranas
Retículo Endoplasmático
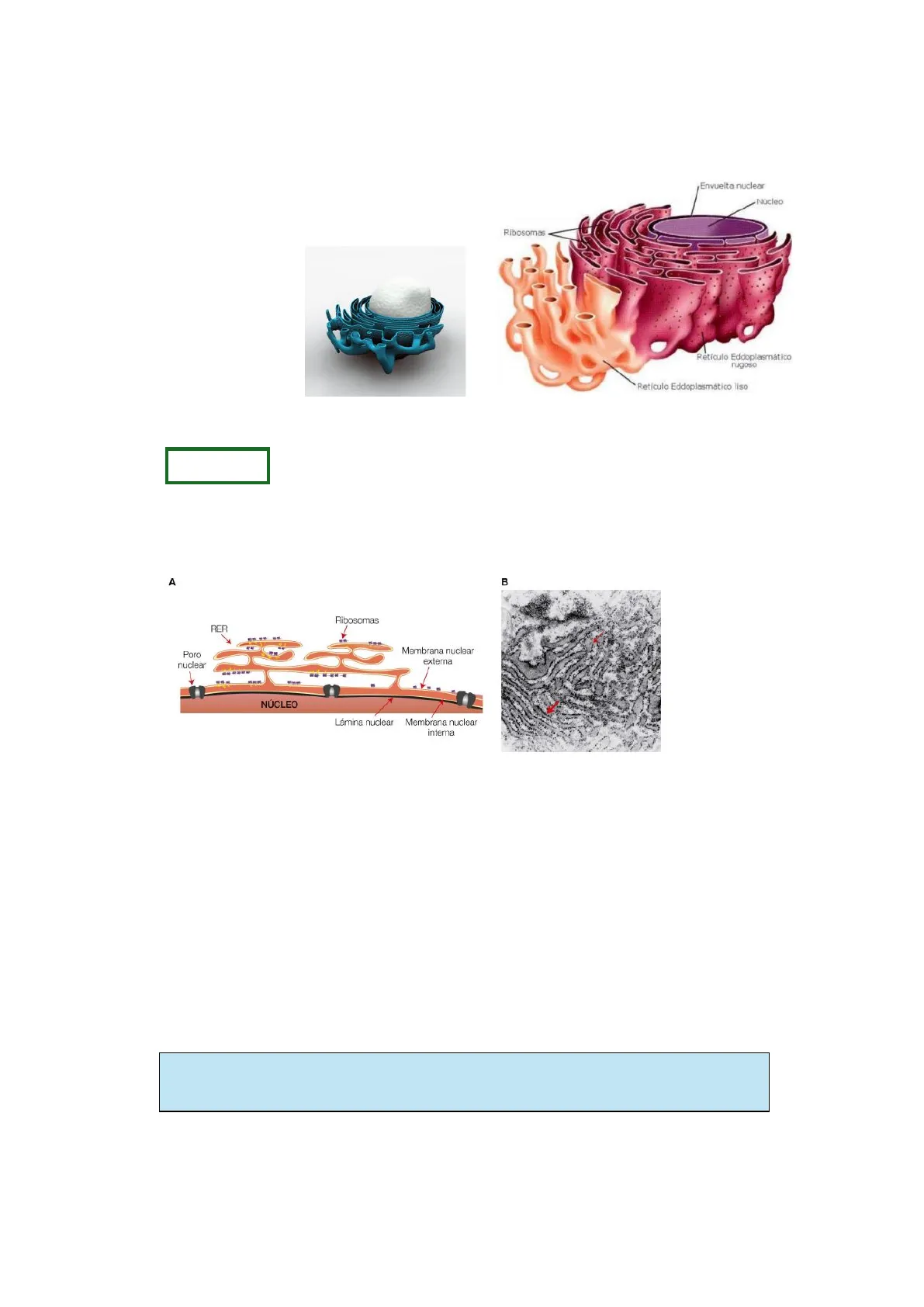
TEMA 4: SISTEMAS DE ENDOMENBRANAS 1. RETÍCULO ENDOPLASMÁTICO El retículo endoplasmático se divide en: · Rugoso (RER) · Liso (REL) Envuelta nuclear Núcleo Ribosomas Retículo Eddoplasmático rugoso Reticulo Eddoplasmático liso
Retículo Endoplasmático Rugoso (RER)
RUGOSO Se trata de una red interconectada de sáculos (o cisternas) aplanados y túbulos membranosos con ribosomas adheridos a su superficie. Estos saculos se conectan con la envoltura nuclear y con el Retículo Endoplasmático Liso.
A B Ribosomas RER Poro nuclear Membrana nuclear externa 1 NÚCLEO Lámina nuclear Membrana nuclear interna
Basofilia Citoplasmática y Tipos
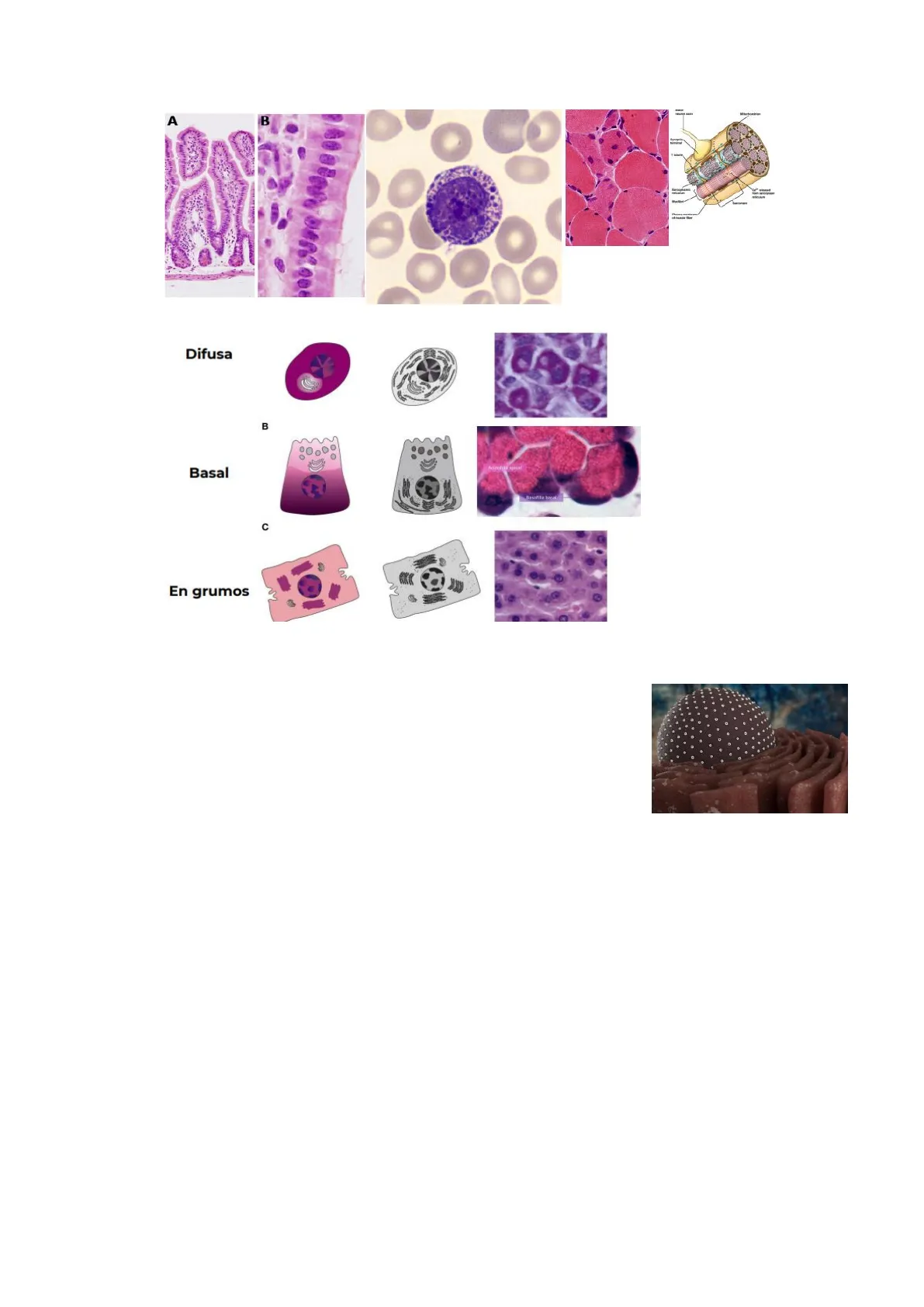
Se encuentra presente en todas las células nucleadas (excepto espermatozoides). Las células con abundante RER presentan una propiedad denominada basofilia citoplasmática, que consiste en la apetencia por los colorantes básicos que presentan ciertas áreas del citoplasma celular. Esta basofilia citoplasmática se corresponde con zonas de abundancia del ARN de los ribosomas libres en el citosol o presentes en el RER. En función a la disposición de este componente basófilo se distinguen tres tipos de basofilia:
- Difusa > prácticamente todo el citoplasma es basófilo. Se observa una zona no basófila correspondiente con el AG (zona Golgi negativa). Ej: células plasmáticas
- Basal > se encuentra en la zona basal de la célula. Ej: células epiteliales
- En grumos > aparece en paquetes o grupos que se corresponden con zonas de acúmulos de RER o ribosomas libres. Ej: hepatocitos y neuronas
Hematoxilina es un colorante básico, que tiñe de azul o morado las estructuras ácidas, como el núcleo de la célula (donde está el ADN, que es ácido).A B Difusa B Basal Acidohis apical c En grumos
Funciones del RER
El RER está involucrado en la síntesis y la distribución de proteínas . Existe una ruta sintetica y secretora de proteínas que pasan del RER al Golgi y luego a la membrana plasmática y al exterior celular . La membrana del RER presenta mayor porcentaje de lípidos y una serie de proteínas exclusivas con el fin de permitir la unión de los ribosomas y la translocación de las proteínas hacia su luz . El interior del RER posee proteínas propias (ej . enzimas relacionadas con la glucosilación de proteínas, peptidasas, chaperonas ... ) .
Traducción de Proteínas en el RER
RUGOSO - TRADUCCIÓN DE PROTEÍNAS Las proteínas celulares pueden traducirse completamente en ribosomas libres presentes en el citosol o comenzar su traducción en ribosomas citosólicos pero continuarla en el RER. Esto marcará el destino de las proteínas, ya que aquellas cuya síntesis pasa por el RER irán destinadas a permanecer en el mismo, en el AG, en lisosomas o peroxisomas, en la membrana plasmática o bien serán secretadas al exterior. Aquellas proteínas que se traducen por completo en el citosol podrán permanecer en este o ser incorporadas después del núcleo, las mitocondrias o los peroxisomas. Como la incorporación de estas proteínas a sus orgánulos diana tiene lugar una vez que se han traducido completamente, la importación es postraduccional. En cambio, losribosomas que se adhieren a la membrana del RER sintetizan las proteínas a la vez que estas se incorporan al RER, por lo que es una importación cotraduccional. Las proteínas que se traducen en el RER poseen un péptido señal de 16 a 30 residuos en su región N-terminal. La traducción y la incorporación simultánea de proteínas en el RER tiene varias etapas:
- La traducción comienza en ribosomas libres. Cuando en el extremo N-terminal de la proteína en formación aparece el péptido señal que indica que la proteína debe ser traducida en el RER, este péptido se reconoce por la partícula reconocedora de la señal (PRS). El péptido señal actúa como secuencia de inicio de la translocación al RER.
- La PRS está formada por una molécula de ARN y seis proteínas y consta de varios dominios (de unión al péptido señal, de parada de la traducción, de unión al ribosoma y de unión a GTP).
- La unión de la PRS al ribosoma provoca una parada en la traducción y causa un cambio de conformación en la PRS que le permite ser reconocida y unirse a su receptor en la membrana del RER . El complejo ribosoma/PRS y su receptor interactúan con un translocador (complejo proteico que permite el paso de la proteína en formación hacia la luz del RER) .
SAP HANA 1 Signal aquerce SRP mapaptor Cytosol Gute ER membrane ATP ER human Tranalocon Translocon (closedti lopeni Cleaved signal sequence ADP - P 6 ATP- ADP . P.+ Hac70 Folded Carbohydrate
- El translocador se abre y continua la síntesis proteica, pero el peptido señal queda anclado en la región interna del translocador mediante interacciones proteicas, por lo que no pasa al interior del RER . La proteína en proceso de síntesis va atravesando el translocador y penetrando hacia el interior del RER . Se produce la hidrólisis de GTP y ocurren cambios conformacionales que liberan a PRS .
- Una peptidasa señal asociada a la cara interna de la membrana, corta el péptido señal de la proteína que se está incorporando al RER.
- Se continúa la traducción de la proteína que quedará en la luz del RER. (Todos los pasos para la foto del 2) CCITOSOL Comienzo de traducción Subunidad pequeña ribosoma ARAm de proteina Subunidad grande ribosoma Péptido señal Receptor del péptido señal (SRP) Membrana del reticulo endoplasmático Receptor para SRP Translocador Reinicio de traducción INTERIOR del RETÍCULO CITOSOL 15 Termina la traducción Membrana del reticulo endoplasmático Peptidasa 50 Liberación de péptido señal Proteína plegada INTERIOR del RETICULO
Localización de Proteínas en el RER
RUGOSO - LOCALIZACIÓN DE PROTEÍNAS La incorporación de proteínas no es común a todas las proteínas que se traducen en el RER; puesto que aquellas que quedan situadas en la membrana del RER poseen señales hidrófobas que les permiten anclarse a la bicapa lipídica. Algunas proteínas, poseen además del péptido señal en el dominio N-terminal, una secuencia de parada de la transferencia en una región interna de la proteína, la cual impide que la proteína continúe translocándose hacia la luz del RER, por lo que la región proteica que permanece por detrás de la secuencia de parada de la transferencia cuando sigue traduciéndose quedará orientada hacia el lado citoplasmático (se genera una proteína anclada a la membrana del RER con su externo N-terminal orientado hacia la luz y el extremo C-terminal hacia el citoplasma). Hay otro tipo de proteínas que se orientan de forma diferente en la membrana. Citoplasma COO- NH; NHẸ Citocromo P450 COO - Receptores asociados a proteína G NH; - Transportador de glucosa GLUT1 - Canales de Ca2+ dependientes de voltaje - Transportadores ABC de moléculas pequeñas - Canal CFTR (CH) - Receptor de LDL - Receptor de insulina - Galactosiltransferasa del Golgi - Sialiltransferasa del Golgi Espacio exoplásmico (luz del RE o aparato de Golgi; espacio extracelular) Elongación
Modificaciones Postraduccionales en el RER
RUGOSO - MODIFICACIONES POSTRADUCCIONALES Las proteínas traducidas en el RER deben adquirir su conformación final para que sean funcionales, lo cual implica un correcto plegamiento de la proteína y una serie de modificaciones postraduccionales en el interior del RER: glucosilación, fragmentación y formación de enlaces disulfuro. Solo las proteínas que pasan por el RER en la célula están glucosiladas. La mayor parte de las proteínas que se traducen en el RER sufren glucosilación (exc. muchas hormonas peptídicas entre otras). La glucosilación comienza en el RER y acaba en el AG. Es importante la glucosilación para que se plieguen correctamente y no se agreguen las proteínas. Rugoso: RE rugoso Filamentos de actina Reticulo Endoplasmático rugoso BER Minograta eldrónico pe ( Reticulo Cidoplasmatico
Retículo Endoplasmático Liso (REL)
LISO Está constituido por un conjunto de estructuras membranosas, esencialmente tubulares, sin ribosomas adosados. Se diferencia del RER tanto en su morfología y composición como en su función. Se encuentra conectado al RER formando una estructura o compartimento en contiguidad de membrana. Diferencias con RER:
- No posee ribosomas
- Los elementos membranosos son túbulos (no sáculos)
- Posee una composición distinta de la membrana (diferentes proteínas y más % lípidos)
- Funciones relacionadas con la síntesis de lípidos y derivados lipídicos
- Abundante solo en células especializadas (células musculares, células secretoras de hormonas esteroideas ... )smooth FER M 500 mm
Funciones del REL
LISO - FUNCIONES En todas las células debe llevarse a cabo la síntesis de fosfolípidos y colesterol para renovar la membrana celular, función realizada por el REL. Hay células especializadas en el organismo en las cuales el REL es muy abundante. En estas células tiene lugar la síntesis de determinados lípidos o derivados lipídicos. La síntesis lipídica se desarrolla en varias fases:
- En el citoplasma dos ácidos grasos unidos a coenzima A se unen al glicerol 3- fosfato, formando el ácido fosfatídico, que se inserta en la membrana del REL.
- Una fosfatasa convierte el ácido fosfatídico en diacilglicerol (DAG), formándose la fosfatidilcolina, la fosfatidiletanolamina y la fosfatidilserina. El fosfatidilinositol se genera a partir del ácido fosfatídico También se sintetizan en el REL el colesterol y la ceramida, que van a ser componentes de las membranas junto con los glicerofosfolípidos. La ceramida dará lugar a glucolípidos y a la esfingomielina en el AG, donde se completará la formación de estos lípidos de membrana. Tanto el colesterol como la esfingomielina formarán parte de las balsas lipídicas, microdominios o rafts.
LIPID RAFT Lipid raft Non-lipid raft Extracellular space glycosphingolipids Intracellular space GPCR cholesterol glycosphingolipid cholesterol G protein nantaine
Síntesis y Distribución de Lípidos
La síntesis de lípidos tiene lugar en la hemimembrana citosólica del RE, de forma que la adición de moléculas lipídicas solo tenga lugar en ese lado de la membrana. Para distribuir posteriormente los lípidos en ambas membranas y establecer la asimetría entran en juego unas enzimas denominadas escramblasas.Las escramblasas catalizan el paso, energéticamente desfavorable, de los grupos polares de los lípidos de un lado al otro de la membrana. Los lípidos sintetizados en el REL se distribuirán por transporte vesicular a los diferentes compartimentos membranosos (RER, AG, lisosomas) y a la membrana plasmática. Para transportarse a las mitocondrias y peroxisomas necesitan proteínas citosólicas intercambiadoras de fosfolípidos.
Acetil-CoA CH2 Enzimas citosólicas CH-CH-CH2 O 0 0 Ácido graso 0=c C=0 O w MMC-S-COA Acetil-CoA-ácido graso + CDP-colina CMP HỌC-CH-CH2 C P Colina C OH OH Glicerol fosfato 3 2 CoA 4 P 2 Colina Fosfatidilcolina Ácido fosfatídico Diacilglicerol OH P 4 8 800000 Escramblasa Aciltransferasas Fosfatasa Fosfotransferasa de colina Colina
Otras Funciones del REL
Contribuye también en la formación de lipoproteínas, ácidos biliares y hormonas esteroideas. Ejemplo: la síntesis de hormonas esteroideas tiene lugar en las células endocrinas especializadas y se desarrolla en pasos sucesivos que implican la acción de enzimas mitocondriales y del REL a partir de colesterol. Se forman así hormonas como la testosterona, el estradiol, el cortisol, la aldosterona, etc. HIPOTÁLAMO la tensión arterial La ADENOHIPÓFISIS ++ + NEUROHIPÓFISIS contrac tiên đối chers ....... ACTH TSH GH ESH LTH Produce Inthe También tiene un papel importante en la degradación de sustancias tóxicas liposolubles (ej. fármacos, alcohol, pesticidas, carcinógenos, compuestos dañinos producidos por el propio metabolismo ... ). Se convierten estas sustancias en productos hidrosolubles que pueden eliminarse de forma más sencilla por el organismo. En los hepatocitos, cuando hay altas dosis de estas sustancias tóxicas hay un REL muy desarrollado que presenta enzimas detoxificantes (ej. Familia de citocromos P450 anclados al lado citosólico de la membrana del REL). Estas enzimas oxidan, hidrolizan o reducen sustancias que tienen que ser modificadas químicamente para reducir su toxicidad. Otras enzimas citoplasmáticas actúan conjugando estas sustancias con determinados grupos químicos (sulfato, acetilo .... ) que facilitan su secreción.
¿Non has encontrado lo que buscabas?
Explora otros temas en la Algor library o crea directamente tus materiales con la IA.
