Canali Ionici del Tessuto Muscolare Cardiaco e Fisiologia Cardiaca
Documento del Prof. Franchi su Canali Ionici del Tessuto Muscolare Cardiaco. Il Pdf, un appunto universitario di Biologia, esplora la fisiologia cardiaca, i canali ionici (HCN, calcio, potassio) e il potenziale d'azione, includendo gli effetti del sistema nervoso parasimpatico e i postulati di Einthoven per l'ECG.
Mostra di più17 pagine


Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Canali Ionici del Tessuto Muscolare Cardiaco
Fisiologia I, Lezione 14, 26/03/2024
Prof. Franchi
CANALI IONICI DEL TESSUTO MUSCOLARE CARDIACO
Il prof inizia la lezione riprendendo alcuni concetti delle lezioni
precedenti.
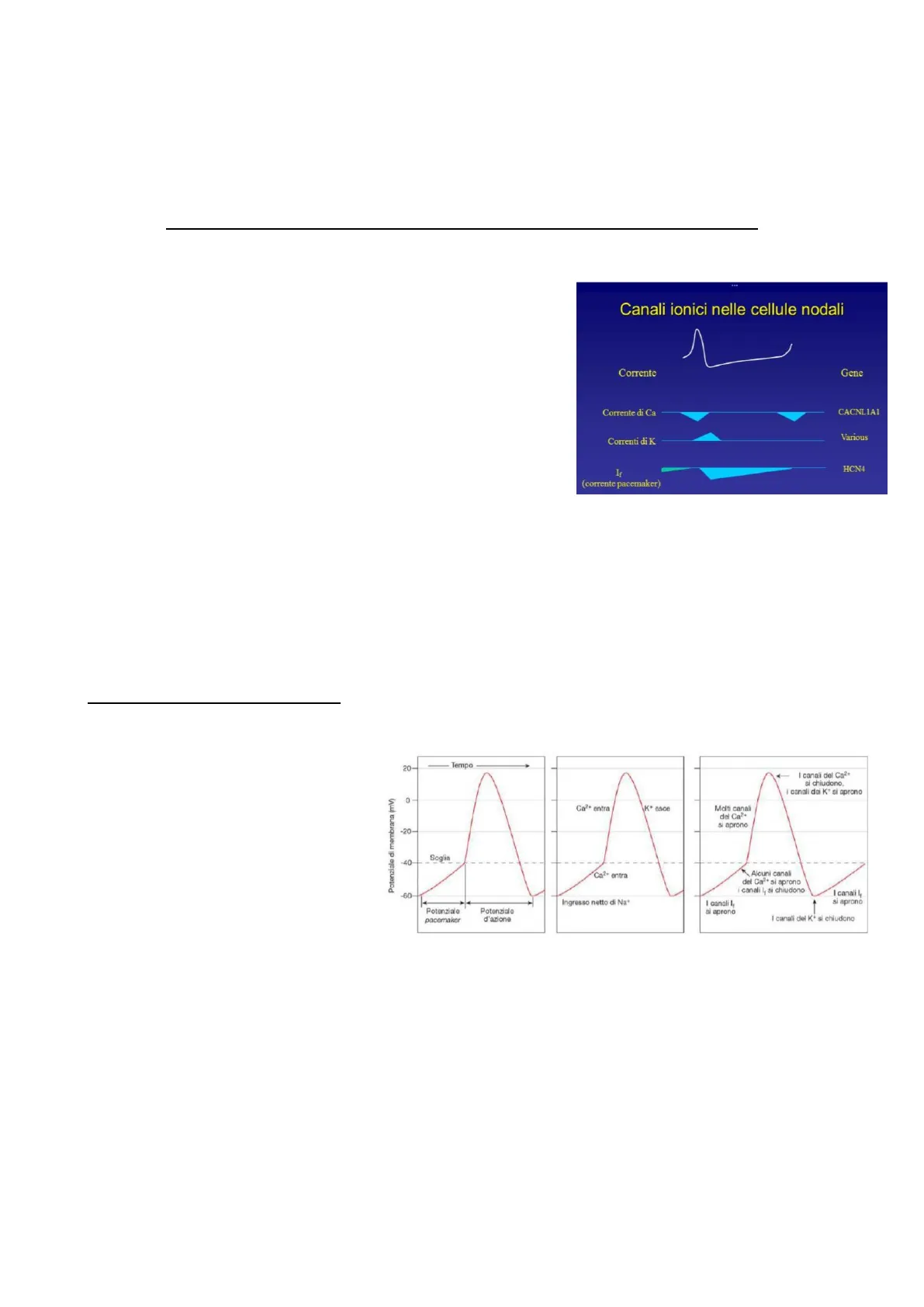
Il primo tracciato in alto rappresenta il potenziale d'azione della
cellula nodale, nella parte sottostante troviamo:
- La corrente calcio i cui picchi si trovano nella parte
discendente e rappresentano una depolarizzazione, - La corrente potassio che rappresenta una corrente varia
- La corrente F rappresentata dai canali HCN
Canali ionici nelle cellule nodali
Corrente Gene
Corrente di Ca CACNL1A1
Correnti di K Various
HCN4
(corrente pacemaker)
I canali HCN hanno la caratteristica di avere una doppia regolazione, questi sono sensibili al voltaggio,
sensibilità elettrica, ma possono essere anche sensibili alla concentrazione di cAMP, sensibilità chimica.
Questi canali si occupano del controllo dell'attività delle cellule nodali in quanto possiedono questa doppia
regolazione.
Canali ionici del nodo seno atriale
Canali ionici del nodo seno atriale:
Il nodo seno atriale è regolato da diversi
meccanismi:
- Canali HCN o corrente pacemaker: il potenziale diventa
gradualmente meno negativo, ho
una depolarizzazione, fino a che
non raggiunge la soglia
innescando un potenziale d'azione (grafico 1)
-20
Soglia
-40
-60
20
- I canali del Ca2+ si chiudono,
i canali doi K* si aprono
Potenziale di membrana (mV)
Ca2+ entra
K* esce
Molti canali
del Ca2+
si aprono
Ca2+ entra
* Alcuni canali
del Ca2+ si aprono
i canali ly si chiudono
I canali I,
si aprono
Ingresso netto di Na*
I canali I,
si aprono
1
I canali del K* si chiudono
Potenziale
pacemaker
Potenziale
d'azione
- Stato funzionale e flusso dei canali ionici, canali calcio (grafico 2)
- Tempo
0
Per cosa differenziano i canali Calcio di tipo T e di tipo L? (La risposta nella registrazione non risulta
comprensibile: quella che segue è un'integrazione del Conti). Dal punto di vista funzionale possiamo
raggruppare i canali calcio in due grandi classi in base alla loro soglia di attivazione: canali ad alta soglia
e canali a bassa soglia. I canali L sono canali ad alta soglia: si attivano a seguito di una marcata
depolarizzazione, ovvero quando il potenziale di membrana raggiunge valori intorno a -20 mV, hanno una
velocità di attivazione minore e sono più lenti. I canali T sono canali a bassa soglia; la loro soglia di
1attivazione è compresa tra valori del potenziale di membrana che vanno da -65 mV a -50 mV, hanno
un'attivazione più rapida; si aprono a valori di potenziale vicini al potenziale di membrana e generano un
ingresso di ioni calcio alquanto fugace.
Quando il potenziale arriva a soglia, circa -40mV, abbiamo una depolarizzazione dovuta all'apertura dei
canali calcio, l'ingresso dei canali calcio determina una fuoriuscita degli ioni K e una conseguente
ripolarizzazione fino a raggiungere il valore di -60mV. Si avrà poi una ulteriore apertura dei canali HCN per
ricominciare il processo e tornare al valore soglia. I canali HCN lasciano passare diversi cationi ma il flusso
principale è rappresentato dagli ioni Na.
L'attivazione dei canali HCN è lenta e avviene a valori di
iperpolarizzazione compresi tra -70 e -50 mV e sono modulati
dalla concentrazione intracellulare di cAMP. Si genera una
corrente definita Corrente Funny generata da ioni Na e altri
cationi. La seconda fase di depolarizzazione é costituita da una
corrente di ioni Ca: i canali si attivano a un potenziale di
membrana di circa -55mV e la depolarizzazione che si crea é
minore rispetto a quella della corrente funny.
Responsabili del Potenziale Pacemaker
RESPONSABILI POTENZIALE PACEMAKER:
- Corrente pacemaker If (funny corrent) di Nat e altri
cationi:
· Lenta attivazione a Vm tra -70 e -50 mV, (valori di Vm
normalmente raggiunti alla fine del PA).
· Modulata da variazioni di AMPc, che modifica l'attivazione
voltaggio dipendente del canale (TAMPc -> attivazione a Vm meno
negativi e viceversa). - Corrente di Ca2+ di tipo T:
· Attivazione a Vm -55 mV, responsabile della seconda fase della
depolarizzazione. - Inattivazione corrente di K+, (IKir = inward rectifier)
normalmente attiva a Vm vicini al potenziale di riposo. (non è la causa
del potenziale di pacemaker, ma la conseguenza.
I canali citati fino ad ora sono responsabili del potenziale pacemaker: canali HCN e canali calcio di tipo T.
Responsabili del Potenziale d'Azione
I responsabili del potenziale d'azione sono i canali calcio di tipo
L, detti anche slow inward, e i canali per il potassio. Si ha
un'apertura dei canali per il calcio che saranno responsabili di
una depolarizzazione con aumento del potenziale d'azione.
Durante la fase di depolarizzazione si ha l'attivazione dei canali
per lo ione K e una successiva ripolarizzazione. In questa fase
non sono presenti ioni Na e la ripolarizzazione é tutta ad opera dei canali K.
RESPONSABILI POTENZIALE D'AZIONE:
- Corrente di Ca2+ di tipo L (Isi = slow inward):
· Attivazione a Vm -40 mV, responsabile della fase di salita del
PA - Corrente K+ (Ikv)
· Attivazione in fase di depolarizzazione, responsabile della
fase di ripolarizzazione del PA
La corrente Na+ voltaggio dipendente nel tessuto nodale è
inattiva al Vm di riposo.
Canali ionici delle cellule del Purkinje
Canali ionici delle cellule del Purkinje:
Le cellule del Purkinje hanno un potenziale d'azione che assomiglia più a quello del miocardio comune che a
quello nodale. Sono presenti tutti i canali sodio e potassio (presenti anche nel miocardio commune). La
differenza tra questi due tessuti é che nelle cellule di Purkinje é presente anche una componente di corrente
calcio la quale determina l'autoeccitabilitá di queste cellule.
La corrente che si crea é molto piccola e molto lenta e va ad incrementare l'attività nodale. L'impulso può
partire anche dalle cellule del Purkinje o può partire un' extrasistolia, che può generare un potenziale
d'azione che fa contrarre l'auricola.
Diffusione del Potenziale d'Azione nel Miocardio
2I fattori che favoriscono la diffusione del potenziale d'azione nel miocardio comune sono:
- Ampiezza del potenziale d'azione, periodo refrattario effettivo ovvero periodo in cui si genera un
potenziale d'azione che non è sufficiente ad eccitare la cellula - Velocità della variazione del potenziale
- Valore potenziale di riposo
- Eccitazione prematura del miocita non completamente
ripolarizzato
T
Tensione muscolare -+
T,
Tensione
isometrica
massima
Tensione
attiva
Tensione
passiva
(b) I muscoli scheletrici se stimolati ripetutamente mostreranno
sommazione e tetano (i potenziali d'azione non sono riportati).
Potentiale di membrana [mV)
Contrazione musco
Pilascurrento mico
Tensione
8
Periodo refrattario
(c) La lunga durata del periodo refrattario nel muscolo cardiaco
previene il tetano.
Tensione
C
Variando la lunghezza alla quale il muscolo è fatto contrarre, si
ottengono valori diversi di tensione. Ponendo questi valori in grafico, in
T
funzione della lunghezza, si ottiene la curva lunghezza-tensione. La
L
tensione effettivamente prodotta ad una determinata lunghezza è la
Lmax
somma della tensione passiva ( tensione che il muscolo sviluppa quando
Lunghezza muscolare -+
viene stirato) e della tensione attiva ( tensione che il muscolo sviluppa quando si contrae in condizioni di
rposo). L'analisi della relazione lunghezza- tensione delle fibre muscolari cardiache permette di
comprendere la successione di eventi contrattili che sono alla
(a) Fibra rapida del muscolo scheletrico: li periodk
refrattario (giallo) e molto breve rispetto al tempo
necessario per lo sviluppo della tensione muscolare.
Tensione
Periodo
refrattario
massima
base del funzionamento cardiaco.
Il muscolo scheletrico lavora per reclutamento. Il reclutamento
0
10
100
0 75 150
Tempo (me)
Stimolo
Tompo (ma)
avviene grazie a uno stato di contrazione tetanica che deriva da
(c) Fibra muscolare cardiaca: Il periodo refrattario dura
quasi quanto la contrazione muscolare
Reco
- Periodo refrattario-+
una serie di contrazioni derivanti da una frequenza specifica detta
frequenza di fusione. Il muscolo cardiaco ha un potenziale
Tensione
90
-Periodo refrattario-+
Potenziale di mombrana |miV)
Potenzialo di mombrana (mv)
d'azione con un lungo periodo di refrattarietà. La curva del
200
250
250
.
Stimolo
Tempo (maj
.
.
potenziale d'azione e quella della tensione possono quindi essere
Fisiologia
ea
wyright 2005. 2000 Cana Barrios Andowsiana
Tempo (ms)
sovrapposte in modo da aumentare la durata del potenziale d'azione cardiaco e quindi del periodo
refrattario assoluto poiché la contrazione cardiaca si sviluppa quando il potenziale d'azione non è ancora
terminato. Durante la maggior parte della contrazioni cardiache le fibre muscolari risultano ineccitabili.
Questo impedisce nel miocardio lo sviluppo di una contrazione tetanica.
La frequenza critica di fusione del muscolo scheletrico e del muscolo cardiaco non sono uguali e differiscono
per la concentrazione del calcio.
Quindi nel muscolo scheletrico abbiamo una contrazione tetanica: questa avviene attraverso una serie di
piccole contrazioni che andranno ad accumulare calcio. È una contrazione che avviene per reclutamento di
unità motorie. Le unità motorie vengono stimolate da un motoneurone che a sua volta va ad attivare anche un
interneurone inibitorio che inibisce lo stesso neurone che lo ha eccitato, inibizione attiva. Le prime unità
motorie ad essere attivate sono anche le prime ad essere inibite.
Meccanismo di Contrazione Cardiaca
3Tutto questo meccanismo nel cuore viene a mancare, il cuore ha una
contrazione massimale.
La contrazione delle fibre cardiache dipende dal calcio che entra durante il
plateau del potenziale d'azione e che promuove il rilascio di calcio dal reticolo
sarcoplasmatico, vengono quindi attivati i canali calcio noti come recettori per
la rianodina. Il rilassamento delle fibre cardiache dipende dalla chiusura di
T-tubule
questi canali e dalla rimozione rapida del calcio citoplasmatico operata
dall'azione simultanea delle pompe calcio dipendenti che riaccumulano il
calcio nel reticolo sarcoplasmatico e allo scambiatore Na/Ca che espelle calcio all'esterno.
NELLA CELLULA MUSCOLALE CARDIACA | SEGNALI CALDO SONO REGOLATI DAI RECETTORI
NELLA CELLULA MUSCOLALE CARDIACA I SEGNALI CALCIO SONO REGOLATI DAI RECETTORI
B-ADRENERGICI
RyR2
CELL MUSCOLARE
CARDIACA
Calcium- induced calcium release (CICR)
VOC di tipo L (recettori di diidropiridine)
Sarcolemma
2
Ca
Mychiaments
Contraction
200 m
PLB = fosfolambano; inibitore di SERCA regalato negativamente da fosforilazione
Tono Vagale Simpatico e Innervazione Cardiaca
Tono vagale simpatico:
Il nodo senoatriale è sotto il controllo costante del sistema nervoso
simpatico e parasimpatico. La frequenza cardiaca normale è il risultato
della contemporanea modulazione positiva del simpatico e negativa del
vago. Il tono vagale prevale sul tono simpatico.
Il prof pone domande sull'anatomia dell'innervazione del cuore. Per
completezza della spiegazione si riporta un breve riassunto di questa.
" il cuore è innervato dal plesso cardiaco alla cui formazione concorrono
fibre parasimpatiche provenienti dal nervo vago e da fibre simpatiche o
(Pre-ganglionic)
orto simpatiche derivate dai gangli del tratto cervicale toracico della
To SA node, AV node.
atrial and ventricular
Cardiac plexus
muscle
catena dell'orto simpatico. I rami cardiaci del vago originano nel collo e
nel torace e si riuniscono in un gruppo superiore, uno medio e uno
inferiore. Tutti i nervi cardiaci del vago e dell'orto simpatico si portano al plesso cardiaco. Dal plesso
cardiaco originano filamenti nervosi che ,accompagnando le arterie coronarie destra e sinistra, si
distribuiscono al cuore. Alcune fibre si portano al nodo senoatriale e nodo atrio ventricolare, oltre al
miocardio atriale e ventricolare e alle pareti dei grossi vasi.
Il ventricolo è innervato unicamente da fibre del simpatico mentre l'atrio è
innervato da fibre del vago, sistema parasimpatico, e da fibre del
simpatico. Il ventricolo, essendo innervato solo dal simpatico, possiede
solo un'inibizione mentre l'atrio avendo doppia innervazione possiede un
doppio controllo, una doppia inibizione.
Si tratta di fibre bianche mieliniche colinergiche.
Stimolazione vagale
Frequenza cardiaca
180
(battiti/min)
120
60
0
7 Hz; 22 s
10 Hz; 20 s
A
Stimolazione simpatica
300
Frequenza cardiaca
(battiti/min)
=240
180
5. 120
20 Hz; 30 s
60
0
B
Thalamus
Medulla
Cervical
sympathetic
ganglia and
nerves
Superior
Vagus nerves
Middle
Inferior
(Stellate ganghon)
(Via superior and inferior
cardiac branches, and
thoracic cardiac branches
of right and left vagus )
Thoracic sympathetic
gangha and
nerves
(Post-ganglionic)
To SA node, AV node,
atrial and ventricular
muscle
Figura 18-2 Variazioni della frequenza cardiaca evocate
dalla stimolazione (barre orizzontali) del nervo vago (A)
e simpatico (B). (Modificato da Warner, H.R. e Cox, A., J Appl
Physiol 17:349, 1962.)
4
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
