Sviluppo embrionale del Zebrafish: segmentazione e gastrulazione
Slide sullo Sviluppo Zebrafish. Il Pdf, utile per studenti universitari di Biologia, descrive lo sviluppo embrionale del Zebrafish, un organismo modello, focalizzandosi sulle fasi di segmentazione e gastrulazione, con diagrammi e immagini.
Mostra di più11 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Sviluppo del pesce zebra
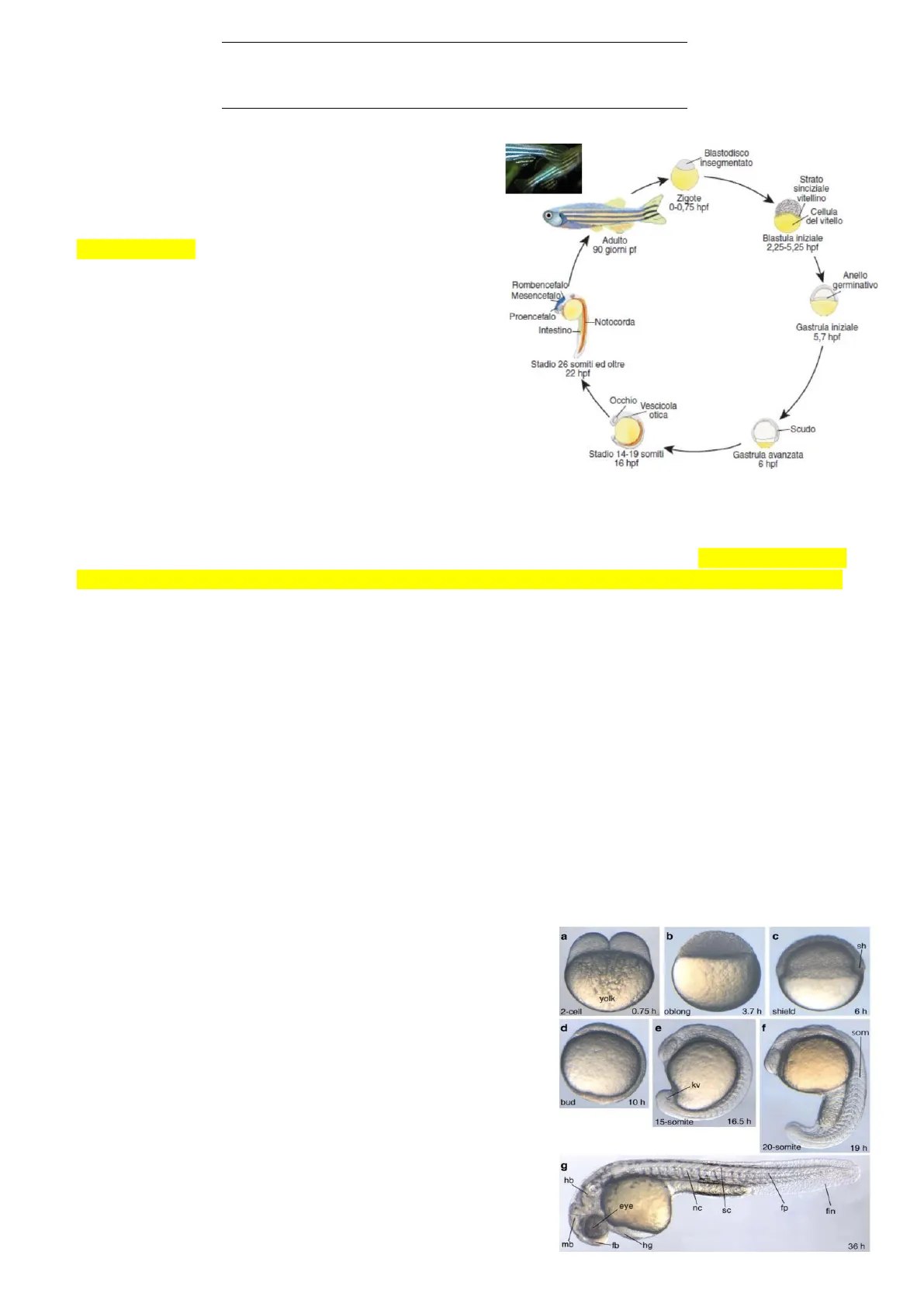
Lo sviluppo dei vertebrati e in particolare del pesce zebra Zebrafish come organismo modello, quindi parleremo innanzitutto dei pesci, poi degli anfibi, dei rettili e infine dei mammiferi. Sviluppo diretto, quindi dopo la fecondazione, attraverso le fasi di: segmentazione, gastrulazione e organogenesi, alla formazione in soli 1/2 giorni della larva, completa di tutte le strutture e che presenta un piano corporeo corrispondente poi a quello dell'adulto. La fase adulta si raggiunge in tempi relativamente brevi di accrescimento, entro 90 giorni ma in condizioni ottimale anche nel giro di 5/6 settimane si ottiene un organismo adulto che a sua volta è in grado di riprodursi ulteriormente. Si tratta di un organismo (l'adulto) di dimensioni relativamente limitate (lunghezza dell'ordine di qualche centimetro).
Blastodisco insegmentato Strato sinciziale vitellino Zigote 0-0,75 hpf Cellula del vitello Adulto 90 giorni pf Blastula iniziale 2,25-5,25 hpf Anello germinativo Rombencefalo Mesencefalo) Proencefalo -Notocorda Gastrula iniziale 5,7 hpf Stadio 26 somiti ed oltre 22 hpf Occhio Vescicola otica Scudo Stadio 14-19 somiti 16 hpf Gastrula avanzata 6 hpf
L'isolamento dei geni è stato svolto soprattutto in organismi invertebrati come poi vedremo in Drosophila e in Nematode Elegans. Nei vertebrati è stato molto più difficile a causa delle dimensioni maggiori e del ciclo vitale più lungo, mentre lo zebrafish è stato in particolare un sistema modello abbastanza unico nel suo genere, proprio per le piccole dimensioni, in ambienti non eccessivamente grandi, con opportuno sistema, è possibile mantenere migliaia di individui con tempi di riproduzione (passaggi da una generazione all'altra) relativamente rapidi. Questo permette di: effettuare degli screening genetici che si basano sull'isolamento di linee mutanti che presentano determinate alterazioni dello sviluppo; isolare i geni responsabili delle mutazioni che producono un determinato fenotipo.
Oltre a queste caratteristiche, l'organismo presenta:
- sviluppo embrionale molto rapido
- una struttura anatomica relativamente semplice, rendendo più facile identificare rispetto ad altri organismi fenotipi mutanti a carico dello sviluppo embrionale
- un genoma relativamente compatto, molto utile per identificare mutazioni che producono alterazioni dello sviluppo, sia anche per creare organismi geneticamente modificati (Transgenici o mutanti)
- Tessuti trasparenti durante tutta la fase embrionale quindi attraverso l'analisi microscopica è possibile seguire bene il comportamento di cellule e tessuti, non soltanto all'esterno ma anche all'interno.
Nella foto al microscopio ottico dell'embrione di Zebrafish in varie fasi dello sviluppo. Ad esempio, in g) la struttura bastoncellare (orizzontale) che si vede in trasparenza è la notocorda, nonostante sia localizzata in posizione centrale nell'embrione; e vedete qui come in questo stadio larvale, con una chiara polarità antero- posteriore, sono presenti già i rudimenti delle strutture dell'adulto tutti quanti formati si arriva all'incirca in 1/2 giorni di sviluppo. Intanto ci occupiamo delle fasi precoci dello sviluppo, quindi segmentazione e gastrulazione, e seguendo la sequenza di analisi che abbiamo già effettuato con gli echinodermi ed anche con i cordati non vertebrati, partiamo dall'analisi delle caratteristiche dell'uovo.
a b C sh yolk 2-cell 0.75 h oblong 3.7 h shield 6 h d e f som .kv bud 10 h 15-somite 16.5 h 20-somite 19 h g hb eye nc SC fp fin mb fb `hg 36 h
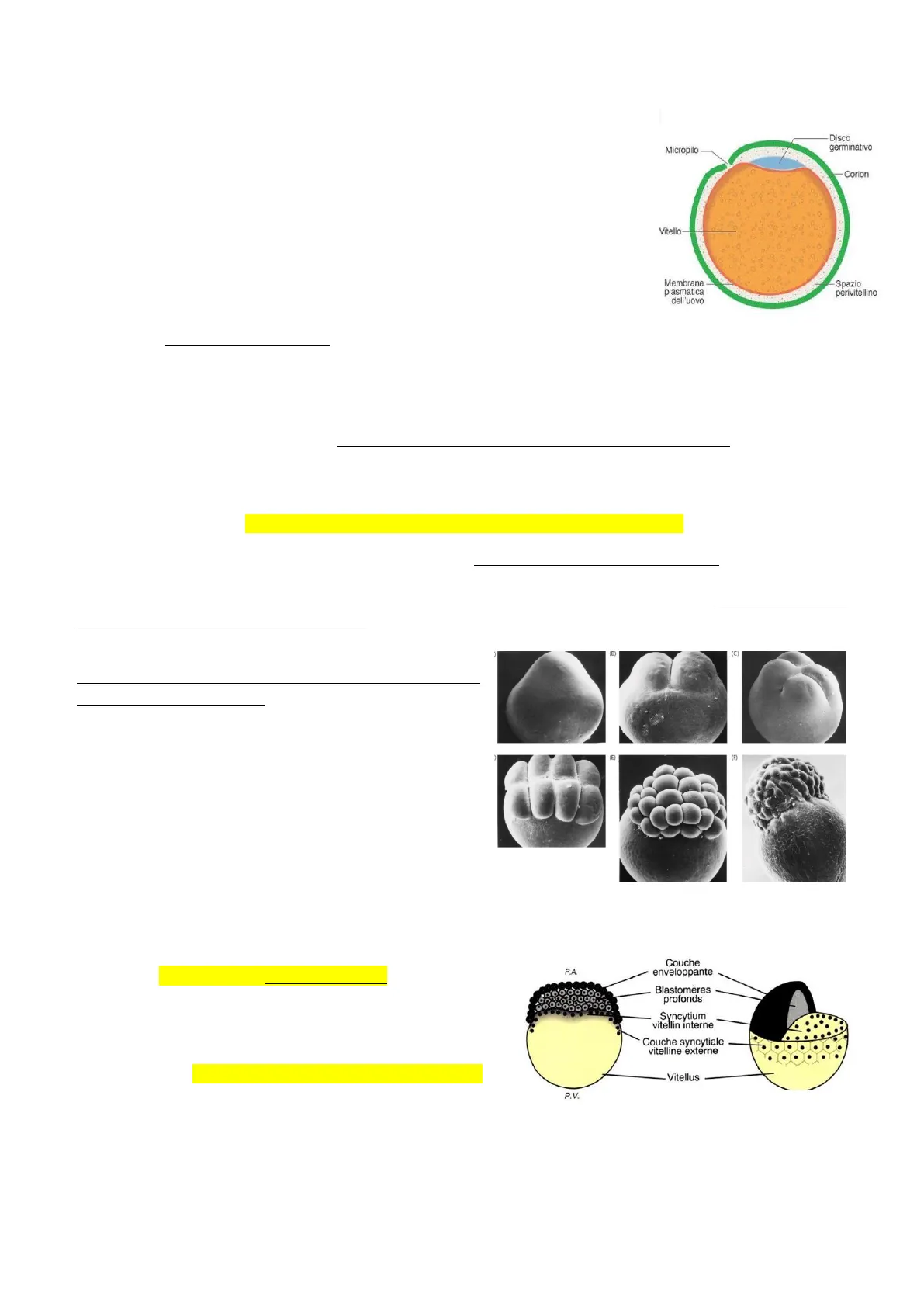
Caratteristiche dell'uovo
- Caratteristiche dell'uovo: Uovo telolecitico, quindi ci troviamo per la prima volta in un organismo che presenta un uovo con elevate quantità di vitello. In pratica tutta la regione centrale e la regione dell'emisfero vegetativo dell'uovo e quindi poi dello zigote, sono molto pieni di vitello così da non andare incontro a divisioni di segmentazione, mentre al polo animale c'è una sottile regione discoidale che prende il nome di disco germinativo dove il tuorlo non è presente. Perciò le divisioni di segmentazione daranno luogo ad una segmentazione di tipo meroblastico, cioè parziale (con piani di segmentazione che interessano solamente la regione del polo animale, quindi il disco germinativo) e discoidale, perché la regione che si viene a segmentare ha appunto questa forma discoidale, e produce quello che viene chiamato un blastoderma discoidale.
Disco germinativo Micropilo Corion Vitello Membrana plasmatica dell'uovo Spazio perivitellino
Segmentazione
- SEGMENTAZIONE È di tipo meroblastica discoidale: le prime divisioni sono divisioni di tipo VERTICALE che dividono il blastoderma discoidale in una serie di cellule che sono tutte organizzate in un monostrato, quindi tutte sullo stesso piano al di sopra del tuorlo. In questa fase le cellule mantengono ancora dei collegamenti con il tuorlo sottostante, ovvero le membrane cellulari separano fra di loro le cellule ma non si chiudono completamente al di sopra del tuorlo; quindi, il citoplasma della cellula e il tuorlo inizialmente sono collegati. Le divisioni di tipo meroblastiche discoidali continuano fino allo stadio di circa 32 cellule. A questo punto il blastoderma è costituito da un monostrato di 32 blastomeri disposti tutti quanti allo stesso livello sopra il tuorlo. In questa fase cominciano anche a chiudersi le membrane cellulari al di sotto delle cellule, separando quindi il citoplasma da quello del tuorlo sottostante.
A questo punto cominciano ad entrare in gioco anche dei piani di divisione ORIZZONTALI che quindi vanno a stratificare il blastoderma generando più strati di cellule gli uni sovrapposti sugli altri al di sopra del tuorlo, come si può vedere in queste fotografie al microscopio elettronico. Vedete le prime divisioni verticali con angoli ruotati di 90° le une rispetto alle altre che portano alla formazione di un blastoderma di 2,4,8 (immagine B-C-D) poi 16 e 32 cellule tutte disposte allo stesso livello (E). Andando avanti si stratifica il blastoderma e ci saranno cellule più vicine al tuorlo e cellule più distanti. Nella fase dell'ultima immagine (F) il blastoderma si stratifica assumendo una forma a cupola o doom al di sopra del tuorlo, cupola non cava ma contenente all'interno due tipi di cellule:
- Una rappresentata in grigio scuro, più esterna che viene detta periderma, un tessuto transitorio che ha la funzione di rivestimento e protezione, si perderà quando l'embrione formerà il vero tessuto epidermico.
- I blastomeri profondi colorati in grigio chiaro che sono quelli che daranno luogo all'embrione vero e proprio. L'embrione si formerà a partire dai blastomeri profondi.
(B) (C) (E (F) P.A Couche enveloppante Blastomères profonds Syncytium vitellin interne Couche syncytiale vitelline externe - Vitellus P.V.
In questa fase avviene anche la formazione di un'ulteriore popolazione che si viene a creare all'interno del tuorlo. Dentro il tuorlo non è possibile la formazione di membrane cellulari e quindi la presenza di piani di divisioni cellulari, però diversi nuclei possono anche trovarsi all'interno del tuorlo, ed è proprio quello che avviene: le cellule che stanno direttamente al di sopra del tuorlo vanno incontro a lisi e riversano all'interno del tuorlo il proprio citoplasma e il proprio nucleo; quindi, si forma una popolazione di nuclei che condividono lo stesso ambiente, formano un sincizio vitellino, (A) Enveloping layer (EVL) Blastoderm Deep cells Internal yolk syncytial layer (iYSL) Yolk syncytial nuclei External yolk - syncytial layer (eYSL) Microtubules Questa è la prima fase di gastrulazione, in cui le cellule profonde (deep - Yolk cell cells) sono circondate dallo strato di rivestimento (EVL). La superficie animale della cellula del tuorlo è appiattita e contiene i nuclei dello strato sinciziale del tuorlo (YSL). I microtubuli si estendono per tutto il citoplasma ricco di tuorlo. I nuclei cellulari derivano dalle cellule situate al margine del blastoderma, le quali hanno liberato i loro nuclei nel citoplasma ricco di tuorlo. Di fatto questi nuclei che cadono dalle cellule sovrastanti che si lisano poi dentro al vitello continuano a dividersi senza formare membrane; quindi, i nuclei nel sincizio vitellino aumentano di numero. In quest'immagine vedete rappresentata E questa fase di formazione del sincizio vitellino, in cui la cellula in A (indicata dalla freccia) ad un certo punto si lisa in B, riversa il suo contenuto all'interno del tuorlo della regione sottostante, ed in D si vedono i nuclei presenti. Chiaramente questo avviene su più cellule le une accanto alle altre, e alla fine si crea questa popolazione di nuclei all'interno del vitello che non sono, al contrario delle cellule del blastoderma, separati da membrane cellulari. In più dalla foto al microscopio ottico si vede molto bene la regione tra il blastoderma cellularizzato (la parte di puntini bianchi della foto) e sotto si vede questo sincizio nucleare con i nuclei che condividono tutti lo stesso citoplasma non essendo separati da membrane.
Mappa presuntiva delle cellule profonde
- Mappa presuntiva delle cellule profonde Nell'immagine è mostrata la mappa presuntiva delle cellule profonde dopo che è cessato il rimescolamento delle cellule. Nello zebrafish, in virtù di questa particolare modalità di segmentazione meroblastica, in cui tutte le cellule si formano soltanto al polo animale dello zigote, non abbiamo la formazione di un blastocele; ciò nonostante, questo stadio di blastoderma viene considerato l'equivalente di uno stadio di blastula, ed è lo stadio al quale è stata fatta un'analisi molto dettagliata della mappa dei territori presuntivi. Quindi si è cercato di capire quali porzioni di questo blastoderma vanno a contribuire ai diversi foglietti embrionali: endodermico, mesodermico ed ectodermico.
Qui viene mostrata una visione complessiva della mappa dei territori presuntivi, che ricorda nei suoi due aspetti principali la situazione già vista a livello dell'anfiosso e dell'ascidia. Innanzitutto, i territori dell'ectoderma (blu) del mesoderma (rosso) e dell'endoderma (giallo) sono presenti a livelli diversi dell'asse animale-vegetativo, con l'ectoderma più vicino al polo animale, l'endoderma più distante e il mesoderma racchiuso fra gli altri due territori. Questa mappa è un'organizzazione spaziale con forti analogie con quelle già viste nei cordati non vertebrati.
Naso occhio Epidermide Encefalo Ectoderma Cresta neurale Midollo spinale Mesoderma Muscolatura di origine somitica Ventrale Sangue Pronefro Pinne Cuore Testa Notocorda Muscolatura Dorsale Tubo digerente Fegato Faringe Endoderma Margine del blastoderma Cellula del tuorlo Polo vegetativo B 0
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
