Apparato Digerente: eccitabilità intestinale, peristalsi e ormoni
Documento del Prof. Franchi sull'Apparato Digerente. Il Pdf, un approfondimento di Biologia a livello universitario, esplora l'eccitabilità dell'intestino, i movimenti di peristalsi e segmentazione, lo svuotamento gastrico e il ruolo degli ormoni gastrointestinali come gastrina, colecistochinina e secretina.
Mostra di più18 pagine

Visualizza gratis il Pdf completo
Registrati per accedere all’intero documento e trasformarlo con l’AI.
Anteprima
Eccitabilità dell'intestino
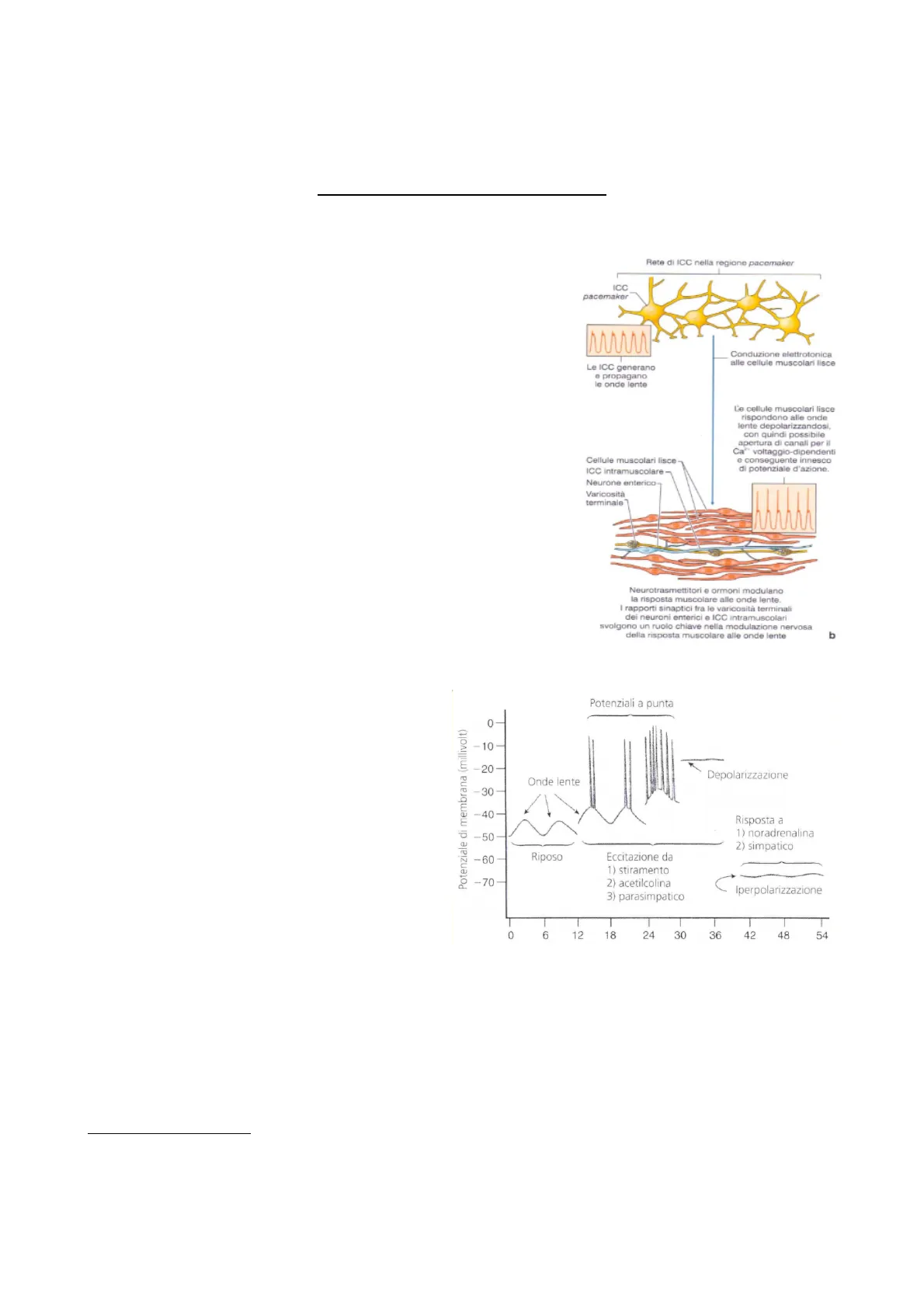
Confrontiamo l'eccitabilità dell'intestino con quella del cuore. Entrambi sono apparati vegetativi, quindi indipendenti dall'attività del SNC. Entrambi presentano delle cellule pacemaker. Qual è la differenza? La differenza è notevole. Nel cuore abbiamo visto che queste cellule sono segregate in dei nuclei (ricorda la rilegatura di Stannius). Questi nuclei hanno un'autoeccitabilità di riposo, non hanno quindi un potenziale d'azione di riposo, ma, come arriva il potenziale d'azione, si autodepolarizzano fino alla soglia di 15mV. Fanno così partire uno spike che si diffonde, secondo le regole che abbiamo visto a suo tempo, a tutto il miocardio, eccitando tutte le cellule in modo sincrono. Inoltre, abbiamo visto che la regolazione è intrinseca, oppure estrinseca, legata all'attività del simpatico o del parasimpatico, che si equivalgono a livello del nucleo, mentre a livello del ventricolo invece prevale il simpatico. Nell'intestino le cellule pacemaker sono, invece, diffuse per tutta la lunghezza, dallo stomaco al retto, come una rete di cellule che presentano una contiguità stretta con le vescicole del sistema vegetativo, sia simpatico che parasimpatico. Inoltre, sono anche in connessione diretta con le cellule della muscolatura liscia. Queste cellule pacemaker, però, generano delle onde lente che si diffondono a tutta la muscolatura. Le onde lente sono onde sottosoglia di per sé, che non generano un potenziale d'azione ma sono sottoposte ad un controllo eccitatorio che aumenta l'ampiezza dell'onda facendo comparire un potenziale d'azione. Ciò avviene dopo il raggiungimento della soglia, che è a 30-40 mV. Queste onde lente possono essere trasmesse ai neuroni di Meissner, che però non sono sufficienti a indurre la contrazione della cellula muscolare; quando arrivano a soglia, c'è un potenziale d'azione che si inscrive sul potenziale a onda lenta. Rete di ICC nella regione pacemaker ICC pacemaker Conduzione elettrotonica alle cellule muscolari lisce Le ICC generano @ propagano le onde lente Cellule muscolari lisce ICC Intramuscolare Neurone enterico Varicosità terminale Neurotrasmettitori e ormoni modulano la risposta muscolare alle onde lente. I rapporti sinaptici fra le varicosità terminali dei neuroni enterici e ICC intramuscolari svolgono un ruolo chiave nella modulazione nervosa della risposta muscolare alle onde lente b Potenziali a punta 0 Potenziale di membrana (millivolt) -10- -20 Depolarizzazione -30- -40- Risposta a 1) noradrenalina -50 2) simpatico -60 Riposo Eccitazione da 1) stiramento -70- 2) acetilcolina Iperpolarizzazione 3) parasimpatico 0 6 12 18 24 30 36 42 48 54 La differenza dal cuore è che il cuore funziona da pompa, qui invece deve far progredire il contenuto intestinale di pasti diversi, che si mescola nel colon. L'intestino ha un'attività motoria diversa nelle fasi digestive, cioè immediatamente dopo un pasto e un po' di tempo dopo. Della motricità prandiale, cioè che si manifesta durante l'assunzione di cibo, ci sono un movimento di segmentazione e uno di peristalsi. Entrambi sono di natura riflessa, cioè, nascono dalla distensione della parete, quindi dall'attivazione di meccanocettori, neuroni sensoriali di I ordine intrinseci. Nella lezione scorsa abbiamo fatto una classificazione delle cellule, distinguendole in neuroni sensoriali primari, cioè intrinseci, interneuroni sensitivi che attivano solo localmente la muscolatura, rimanendo in sede 1 L'e cellule muscolari lisce rispondono alle onde lente depolarizzandosi, con quindi possibile apertura di canali per il Ca" voltaggio-dipendenti e conseguente innesco di potenziale d'azione. Onde lentenell'intestino, ed estrinseci, efferenti di I ordine con il corpo cellulare nei gangli dei nervi cranici o della rete dorsale.
Movimento di peristalsi
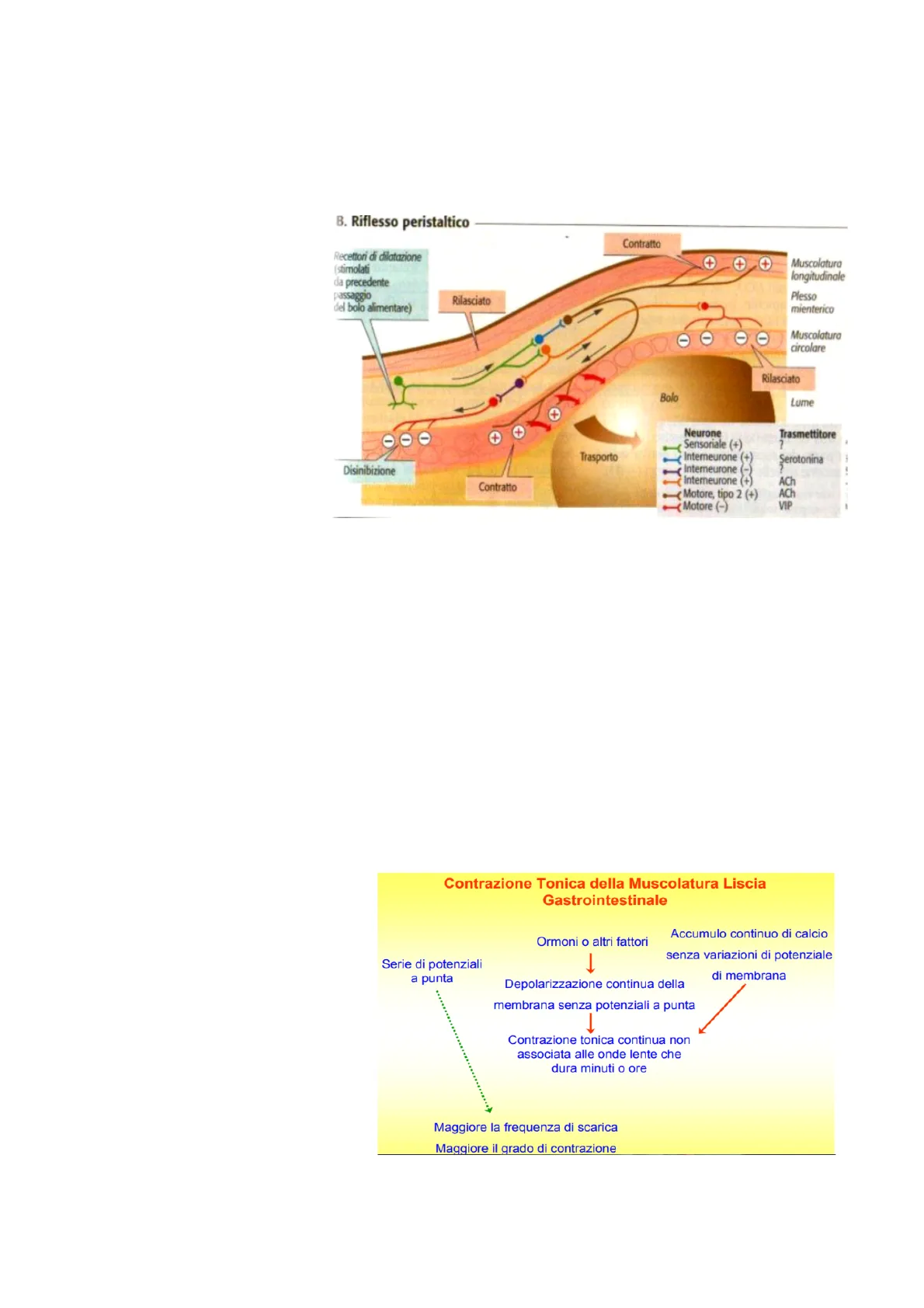
A seguito della distensione della parete, i meccanocettori iniziano il movimento di peristalsi. Abbiamo un movimento ricettivo e uno propulsivo. Nella fase ricettiva della peristalsi c'è contrazione della muscolatura longitudinale, mentre in quella propulsiva della circolare. Nascono da un riflesso intrinseco (interneurone verde). Mostra una figura che vale la pena di saper leggere. B. Riflesso peristaltico Contratto Recettori di dilatazione + Muscolatura (stimolati longitudinale da precedente passaggio Rilasciato Plesso mienterico del bolo alimentare) Muscolatura circolare Rilasciato Bolo Lume Neurone Sensoriale (+) Trasmettitore ? Trasporto Interneurone (+) Interneurone Serotonina ? Disinibizione Interneurone (+) ACh Contratto «Motore, tipo 2 (+) ACh Motore (-) VIP È fondamentale avere contenuto intestinale per il funzionamento ottimale dell'intestino. La distensione della parete attiva il neurone afferente intrinseco il quale va ad attivare motoneuroni eccitatori o inibitori a seconda se è stata la muscolatura longitudinale o circolare.
Movimento di segmentazione
È l'altro movimento riflesso, che nasce anch'esso dalla contrazione indotta dalla distensione della parete. Tuttavia, ha funzioni diverse. La peristalsi è propulsiva, mentre i movimenti di segmentazione sono di rimescolamento. Siccome i processi digestivi riguardano il contatto tra enzimi e substrato, rimescolare significa migliorare il rapporto che c'è tra enzimi e contenuto intestinale, quindi a favorire sia processi di secrezione che di assorbimento. Domanda: Se entrambi sono movimenti riflessi ... (come fa l'intestino ad innescarne uno piuttosto che l'altro)? Risposta: Non sappiamo ancora perché si innesca uno o l'altro perché raramente sono processi isolati, spesso, come da figura, si propagano e sono presenti in un ampio tratto dell'intestino. Questi sono movimenti fasici. Abbiamo Contrazione Tonica della Muscolatura Liscia Gastrointestinale due tipi di movimenti tonici. O quando c'è un'iperattivazione dei neuroni Ormoni o altri fattori Accumulo continuo di calcio eccitatori, in cui si ha un rilasciamento e 1 quindi nessuna attività motoria; oppure Serie di potenziali senza variazioni di potenziale a punta Depolarizzazione continua della di membrana quando c'è un movimento "di massa" originato dai motoneuroni eccitatori che membrana senza potenziali a punta 1 induce una depolarizzazione persistente Contrazione tonica continua non associata alle onde lente che dura minuti o ore senza presenza di spike, cioè onde lente. È uno stato di eccitabilità delle cellule muscolari che non presenta lo spike. C'è una depolarizzazione sostenuta e Maggiore la frequenza di scarica costante che è sostenuta soprattutto dai Maggiore il grado di contrazione motoneuroni eccitatori. C'è anche uno stato di decontrazione in assenza di potenziale d'azione, come uno stato di iperpolarizzazione della membrana delle cellule muscolari. Questo provoca un rilasciamento della parete 2intestinale. Di solito questo rilasciamento si ha, come vedremo, per un'attività persistente e continua dei motoneuroni inibitori. Abbiamo quindi 4 tipi di stato motorio dell'intestino. Due moticità fasiche e due toniche che seguono ad un'attivazione persistente dei motoneuroni e di un rilasciamento che in clinica chiamiamo ileo paralitico, che è uno stato appunto paralitico, cioè non si genera movimento perché c'è uno stato di attivazione dei neuroni inibitori. Ricorda la distinzione tra neuroni inibitori ed eccitatori. Domanda: Ma quindi l'ileo si ha solo in depolarizzazione consistente o in entrambi i casi? Risposta: No è dato da iperpolarizzazione, cioè assenza di contrazione.
Sfinteri
Normalmente gli sfinteri presentano un tono miogeno. La loro apertura o rilasciamento, si ha dall'attivazione dei motoneuroni inibitori. In condizioni di chiusura c'è una prevalenza del tono miogeno e scarsa attività dei motoneuroni inibitori. Quando il motoneurone inibitorio aumenta l'attività si produce un'apertura degli sfinteri. Questo vale per tutti gli sfinteri intestinali.
Complesso motorio migrante
Abbiamo visto questi 4 moti in fase prandiale. Adesso vediamo un'attività in fase interprandiale. Si ha un'attività motoria definita complesso motorio migrante. C'è uno schema di movimento che parte dallo stomaco e si diffonde a tutto l'intestino tenue. Distinguiamo 3 fasi del complesso migrante: La prima fase è una fase di silenzio, in cui non c'è nessuna attività motoria, che dura circa 40 minuti-1 ora. L'assenza di attività è data da un ileo fisiologico, cioè un'iperattività dei neuroni inibitori. Dopo 1 ora circa dall'insorgenza del silenzio (ileo-fisiologico), c'è una seconda fase che presenta di spikes e contrazione irregolare e dura circa 30 minuti. Successivamente la terza fase è di circa 15 minuti, data da una sequenza di contrazioni regolari della muscolatura originata nel punto di passaggio tra il corpo dello stomaco e l'antro. Le fasi del Complesso Motorio Migrante (CMM) si verificano sia durante la veglia che durante il sonno Fase I assenza di attività contrattile: durata ca.40-60 min Aumento [Motilina] plasmatica stimola l'insorgenza dei cicli di CMM Il pasto annulla il ciclo 1 ciclo/90-100 minuti Velocità: 2-8 cm/min Fase III Fase II Contrazioni regolari: attività massima (durata ca. 15 min) contrazioni irregolari: 50% dell'attività massima (durata ca.30 min) 3Si ha un ciclo ogni 100 minuti la cui velocità va dai 2 agli 8 cm/min. (Se dite 5 va bene). La funzione è di mandare avanti e ripulire l'intestino tenue. Il complesso viene interrotto dall'assunzione di un pasto, da due ormoni che eccitano l'attività motoria dello stomaco: gastrina e colecistochinina CCk, prodotti da cellule del duodeno. Sono i segni che il cibo è arrivato nello stomaco e inizia l'attività digestiva. Questa secrezione interrompe il complesso migrante e inizia l'attività tipica di segmentazione e peristalsi. Se registrate con un elettrodo a vari livelli dall'antro dello stomaco, lungo il duodeno vedete l'attività che prima è scarsa, poi irregolare e infine regolare e come si distribuisce in maniera costante per tutto l'intestino. Il complesso migrante è un pattern motorio che si presenta a digiuno con le stesse caratteristiche di fase in tutto l'intestino tenue. Motilità interdigestiva: Il Complesso Motorio Migrante (CMM) Origine del CMM nello stomaco distale (contrazioni da fame) o nel duodeno Aumento della contrazione dello sfintere esofageo inferiore Aumento della secrezione gastrica Aumento della secrezione pancreatica 1 Fronte di attività che migra dal digiuno all'ileo 1 Propulsione peristaltica del contenuto luminale in direzione aborale Pulizia del tratto gastrointestinale Fronte di attività del complesso motorio migrante Trasduttori di pressione الاالسليلىليلى Duodeno Digiuno C 10 15 20 25 30 Tempo (min) Figura 56.24 Complesso motorio migrante rilevato da trasduttori di pressione posti nell'antro gastrico e a vari livelli dell'intestino tenue (modificata da Wood, 2003).
Deglutizione
Iniziamo la descrizione dei movimenti dell'albero digerente a partire dal più encefalico: la deglutizione. Cos'è la deglutizione? Molti insistono a dire che la deglutizione è una catena di riflessi ma c'è un centro della deglutizione, il centro pontino vicino al nucleo ambiguo, che la coordina. Si vede grazie ad elettrodi lo stimolo di movimenti che sono molto simili alla deglutizione, questa è l'unica dimostrazione. Si divide in 3 parti: fase volontaria, faringea ed esofagea. Il processo della deglutizione è complicato perché la faringe assolve alla funzione della deglutizione solo per pochissimo tempo, mentre è coinvolta nella respirazione. È importante quindi che la funzione respiratoria non venga compromessa dalla funzione della deglutizione. Si divide in tre fasi: fase volontaria che inizia il processo fase faringea, involontaria (dalla faringe all'esofago) fase esofagea (dall'esofago allo stomaco) Quando il bolo è pronto per essere deglutito esso è spinto volontariamente indietro con movimenti della lingua. Da questo punto in poi la deglutizione diviene automatica e non può essere bloccata. La fase volontaria orale inizia quando il bolo viene spinto verso la parete posteriore della faringe. Lì si ferma e inizia la fase faringea, involontaria come quella esofagea. 4
Non hai trovato quello che cercavi?
Esplora altri argomenti nella Algor library o crea direttamente i tuoi materiali con l’AI.
